- Úvod
- Metody
- Data o rozšíření a klasifikace biomů
- Druhové složení mezi biomy
- Údaje o vlastnostech
- Měření hyperobjemů znaků
- Funkční odlišnost a rozšířenost
- Funkční prostor a podobnost biomů
- Výsledky
- Diskuse
- Knowledge Shortfalls
- Společné vzorce v rámci biomů
- Srovnání mezi biomy
- Závěry
- Dostupnost dat
- Příspěvky autorů
- Financování
- Prohlášení o střetu zájmů
- Poděkování
- Přídavné materiály
Úvod
Ekologové a biogeografové řadí biodiverzitu suchozemských rostlin do klimaticky podmíněných biomů, jejichž fyziognomie je charakterizována růstovými formami a funkčními znaky dominantních druhů (Moncrieff et al., 2016). Koncept biomů má skutečně dlouhou historii sahající až ke Carlu Ludwigu Willdenovi a Alexandru von Humboldtovi. Willdenow si uvědomoval, že podobná podnebí podporují podobné vegetační formy, a Humboldt pozoroval rozšířenou souvislost mezi rozšířením rostlin, fyziognomií a faktory prostředí. Celkově koncept biomů odráží předpoklad, že podobné tlaky prostředí vybírají druhy s podobnými funkčními vlastnostmi nezávisle na jejich evoluční historii. Současně jsou dochované biomy Země do jisté míry fylogeneticky odlišné, přičemž mnoho/většina charakteristických druhů pochází ze specifických linií, které vykazují klíčové adaptace nejen na klimatický výběr, ale i na další tlaky, jako je oheň nebo megaherbivoři (Woodward et al., 2004; Pennington et al., 2006; Donoghue a Edwards, 2014). Vzhledem k tomu, že biomy představují velkoplošné zákonitosti v zemské vegetaci, je pochopení funkčních rozdílů mezi biomy kriticky důležité pro modelování globálního cyklu uhlíku a fungování zemského systému, včetně reakcí na antropogenní globální změny (Bonan et al., 2012; van Bodegom et al., 2014; Xia et al., 2015).
V posledních dvou desetiletích došlo k rychlému nárůstu porozumění funkčnímu rozměru biodiverzity rostlin v kontinentálním měřítku (Swenson et al., 2012; Lamanna et al., 2014; Šímová et al., 2018). Rozsáhlé úsilí o syntézu a analýzu dat definovalo omezený prostor funkčních znaků zahrnující variabilitu v celkové velikosti rostlin, ekonomice listů a hydraulice, známý jako spektrum funkčních znaků (Wright et al., 2004; Díaz et al., 2016). Spektra funkčních znaků definují fyziologické a ekologické kompromisy, které určují životní strategie rostlin (Westoby, 1998; Reich et al., 2003; Craine, 2009) a jejich vliv na sestavování společenstev (Kraft et al., 2008), funkci ekosystémů (Lavorel a Garnier, 2002; Garnier et al., 2004; Kerkhoff et al., 2005; Cornwell et al., 2008), a dokonce i rychlost evoluce (Smith a Donoghue, 2008; Smith a Beaulieu, 2009). Obecnost a užitečnost spekter funkčních znaků rostlin uspíšila jejich začlenění do modelů globálního rozšíření vegetace (van Bodegom et al., 2014), biogeochemického koloběhu (Bonan et al., 2012) a ekosystémových služeb (Díaz et al., 2007; Cadotte et al., 2011; Lavorel et al., 2011). Jen málo studií však zkoumalo komplexní měření funkční diverzity v měřítku biomů napříč dominantními i podřízenými druhy a formami života.
Existují dva protichůdné soubory předpovědí o tom, jak se funkční diverzita liší mezi klimaticky a fyziognomicky odlišnými biomy. Na jedné straně, pokud biodiverzita biomů odráží variabilitu dostupného ekologického prostoru niky, měly by méně taxonomicky rozmanité biomy představovat menší, do značné míry vnořenou podmnožinu funkčního prostoru, který zaujímají rozmanitější biomy. Na druhou stranu vzhledem ke globální povaze základních kompromisů ve struktuře a funkci rostlin a podobným selekčním tlakům působícím podobně napříč druhy mohou společenstva obývající různá prostředí ve skutečnosti sdílet podobné oblasti prostoru znaků (Reich et al., 2003; Wright et al., 2004; Díaz et al., 2016). Nejnovější studie potvrzují, že vztahy mezi klimatem a funkční a taxonomickou diverzitou (tj. druhovou bohatostí) jsou komplexní a závislé na měřítku. V některých případech funkční diverzita úzce sleduje klimatické gradienty, s nižší funkční diverzitou v proměnlivějších a extrémnějších podmínkách (Swenson et al., 2012; de la Riva et al., 2018), a objem funkčního prostoru v místních společenstvech se rozšiřuje s rostoucí taxonomickou bohatostí (Lamanna et al., 2014; Li et al., 2018). Analýzy druhových fondů v regionálním měřítku však naznačují, že velké změny v druhové bohatosti mohou být spojeny s minimálními dopady na funkční diverzitu (Lamanna et al., 2014; Šímová et al., 2015). Reakce taxonomické a funkční diverzity na klima se navíc může mezi hlavními růstovými formami rostlin lišit (Šímová et al., 2018) a relativní diverzita různých růstových forem se může výrazně měnit podél klimatických gradientů, které vymezují různé biomy (Engemann et al., 2016).
Pokrok v analýze funkční diverzity v kontinentálním až globálním měřítku byl částečně omezen nedostatkem dat při katalogizaci taxonomických, distribučních, fylogenetických a funkčních aspektů biodiverzity (Hortal et al., 2015). Navíc i vzhledem k omezeným dostupným údajům jsou se standardizací a integrací rozsáhlých, různorodých souborů dat popisujících geografické rozšíření, funkční znaky a fylogenetické vztahy druhů spojeny značné informatické problémy (Violle et al., 2014). V této práci zkoumáme rozložení funkční diverzity rostlinných druhů napříč biomy Severní a Jižní Ameriky s využitím databáze Botanical Information and Ecology Network (BIEN; Enquist et al., 2016; Maitner et al., 2018), která shromažďuje informace o rozšíření a funkčních znacích >100 000 druhů suchozemských rostlin. Konkrétně zkoumáme variabilitu funkční diverzity a odlišnosti biomů na základě map rozšíření suchozemských druhů rostlin v měřítku buněk sítě 100 × 100 km a komplexního souboru dat šesti funkčních znaků, které odrážejí hlavní osy variability ekologických strategií.
Naše cíle v této studii jsou trojího druhu. Za prvé, abychom poukázali na přetrvávající nedostatky údajů, dokumentujeme rozsah dostupných dat charakterizujících funkční rozmanitost a odlišnost biomů. Za druhé, vzhledem k dostupným údajům charakterizujeme rozložení funkční diverzity v rámci biomů v dominantních i podřízených růstových formách. Tyto analýzy nám umožňují lépe kvantifikovat funkční odlišnost biomu tím, že identifikujeme nejběžnější funkční strategie nejrozšířenějších druhů v jeho rámci. Zatřetí si klademe otázku, zda jsou biomy ve skutečnosti charakterizovány funkčně odlišnými soubory druhů na základě překrývání vícerozměrných hyperobjemů v prostoru funkčních znaků.
Metody
Data o rozšíření a klasifikace biomů
Pro snížení vlivu zkreslení výběru charakteristického pro soubory dat o výskytu sestavené z více zdrojů jsme použili mapy rozšíření BIEN 2.0 pro 88 417 druhů rostlin rozšířených v Severní a Jižní Americe (Goldsmith et al., 2016). Databáze BIEN integruje standardizovaná pozorování rostlin pocházející z herbářových položek a inventarizací vegetačních ploch. Mapy areálů druhů v databázích BIEN byly odhadnuty pomocí jednoho ze tří přístupů v závislosti na dostupných výskytech jednotlivých druhů. U druhů s pouze jedním nebo dvěma výskyty (cca 35 % druhů) byl geografický areál definován jako čtverec o rozloze 75 000 km2 obklopující každý datový bod. U druhů se třemi nebo čtyřmi výskyty byl geografický areál určen pomocí konvexního trupu (cca 15 % druhů). Nakonec byly mapy areálů pro druhy s alespoň pěti výskyty získány pomocí algoritmu modelování rozšíření druhů Maxent s použitím 19 klimatických vrstev jako predikčních proměnných a 19 prostorových vlastních vektorů jako filtrů pro omezení nad předpověďmi modelů (viz Goldsmith et al., 2016 pro další podrobnosti o metodice map rozšíření).
Mapy rostlinných druhů BIEN 2.0 jsme překryli na mapu sítě 100 × 100 km s Lambertovou azimutální rovnoměrnou plošnou projekcí, abychom získali matici přítomnosti/nepřítomnosti druhů pro každou buňku sítě. Na základě klasifikace biomů podle Olsona et al. (2001) jsme každou buňku matice přiřadili k jedné z kategorií biomů. Vzhledem k výpočetním omezením pro následné analýzy jsme některé biomy spojili na základě podobnosti klimatu a vegetace a literatury, abychom získali širokou klasifikaci popsanou v tabulce 1 (s výjimkou vnitrozemských vod, skal a ledu a mangrovů). Ekoregiony Chaco a Caatinga byly klasifikovány jako Xerické lesy a Suché lesy podle Prado a Gibbs (1993), Pennington et al. (2000), Banda et al. (2016), Silva de Miranda et al. (2018).
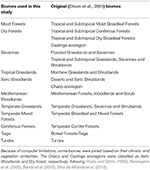
Tabulka 1. Přehled klasifikace biomů přijaté v této studii a ekvivalentní klasifikace biomů (Olson et al., 2001) (s výjimkou vnitrozemských vodních, skalních a ledových a mangrovových porostů).
Druhové složení mezi biomy
Pro pochopení variability v prostoru funkčních znaků mezi biomy jsme nejprve zkoumali rozdíly v druhovém složení rostlin mezi nimi. Na základě seznamu druhů pro každou buňku sítě a biom jsme definovali charakteristické druhy každého biomu jako ty, které mají v daném biomu maximální podíl svého výskytu. Tyto seznamy geograficky převažujících druhů v daném biomu jsme porovnali se seznamem druhů v ostatních biomech. Toto párové srovnání poskytuje jednoduchý způsob, jak odvodit směrový taxonomický překryv mezi biomy (tj. podíl převládajících druhů daného biomu sdílený s jiným biomem). Z 88 417 druhů s dostupnými mapami areálů má 44 899 druhů areály zasahující do více než jednoho biomu a 43 518 druhů je endemických pro určitý biom.
Údaje o vlastnostech
Extrahovali jsme všechny informace o vlastnostech druhů rostlin v Novém světě, které jsou k dispozici v datové sadě BIEN 3.0 (získáno 7. února 2018), čímž jsme získali celkem 80 405 pozorování vlastností na úrovni druhů. Poté jsme vyfiltrovali informace pro šest funkčních znaků: maximální výška rostliny (m), hmotnost semen (mg), hustota dřeva (mg/cm3), specifická listová plocha SLA (cm2/g) a koncentrace fosforu a dusíku v listech na jednotku hmotnosti (mg/g). Po filtraci zůstalo celkem 18 192 pozorování na úrovni druhů. Z nich byla pro další analýzy použita podmnožina 8 820 druhů s mapami areálů i informacemi o znacích.
Pro odhad prostoru funkčních znaků pro každý biom jsme potřebovali kompletní údaje o znacích. Z tohoto důvodu jsme fylogeneticky imputovali chybějící údaje o znacích (Bruggeman et al., 2009; Penone et al., 2014; Swenson, 2014; Swenson et al., 2017) pomocí balíčku R “Rphylopars” v 0.2.9 (Goolsby et al., 2017) a nedávno publikované fylogeneze semenných rostlin od Smitha a Browna (2018) jako výchozí. Fylogenetická imputace je nástroj pro predikci chybějících dat v souborech dat o funkčních znacích založený na předpokladu, že blízce příbuzné druhy mají tendenci mít podobné hodnoty znaků (Swenson, 2014). Použili jsme strom ALLBM (tj. genovou banku a otevřený strom taxonů života s páteří, který poskytli Magallón et al., 2015), protože maximalizoval překryv mezi druhy s dostupnými informacemi o vlastnostech a rozšíření. Po imputaci znaků zůstalo pro další analýzu celkem 7 842 druhů s kompletními informacemi o znacích a mapami rozšíření.
Měření hyperobjemů znaků
Měřili jsme relativní funkční diverzitu biomů výpočtem hyperobjemů znaků ze zásobníků druhů v buňkách biomů. Vzhledem k výpočetním omezením jsme hyperobjemy znaků konstruovali na základě náhodného vzorku 20 % buněk každého biomu. Hyperobjem pro každou buňku sítě byl odhadnut pomocí extrahovaných šesti funkčních znaků a balíčku R “hypervolume” (Blonder et al., 2014, 2018) s použitím Gaussovy metody KDE s výchozím Silvermanovým odhadem šířky pásma. Hmotnost semen, výška a hustota dřeva byly transformovány logaritmicky, zatímco SLA byla transformována kvadraticky. Všechny znaky byly před analýzou škálovány a centrovány. Hyperobjem je uváděn v jednotkách směrodatných odchylek (sd) na mocninu k počtu použitých znaků (tj. sd6).
Funkční odlišnost a rozšířenost
Protože jsou biomy charakterizovány svou dominantní vegetací, zkoumali jsme také geografickou rozšířenost a funkční odlišnost druhů v rámci biomů. Pomocí map rozšíření druhů a informací o funkčních znacích jsme odhadli funkční odlišnost a rozšířenost jednotlivých druhů v každém biomu podle koncepčního rámce funkční vzácnosti Violle et al. (2017). S využitím celého souboru (měřených a imputovaných) znaků jsme nejprve změřili euklidovskou vzdálenost ve standardizovaném prostoru znaků mezi všemi dvojicemi druhů. Poté jsme vypočítali funkční odlišnost (Di) pro každý druh v každém biomu jako průměrnou funkční vzdálenost druhu od N dalších druhů v rámci druhového fondu daného biomu. Di byla v rámci každého biomu škálována mezi 0 a 1, přičemž nižší hodnoty představovaly druhy, které jsou funkčně společné (redundantní), a vyšší hodnoty představovaly druhy, které jsou funkčně odlišné ve srovnání s ostatními druhy v daném biomu. Odhadujeme také geografickou rozšířenost (Wi) druhu v daném biomu, která se měří jako podíl počtu buněk sítě obsazených druhem v rámci biomu na celkovém počtu buněk v daném biomu. Hodnota 1 znamená, že se ohniskový druh vyskytuje ve všech buňkách sítě, které daný biom pokrývá. Funkční odlišnost i geografická rozšířenost byly vypočteny pomocí balíčku R “funrar” (Grenié et al., 2017).
Protože nám chybí komplexní míry dominance založené na lokální početnosti nebo biomase, použili jsme míry Di a Wi, abychom pro každý biom určili “nejběžnější” druhy jako ty, které jsou zároveň geograficky rozšířené (Wi > 0,5) a funkčně podobné (Di < 0,25). Tento poslední práh byl použit, protože třetí kvantil hodnot funkční odlišnosti mezi biomy se pohyboval od 0,2 do 0,3. Pro rozlišení funkčně odlišných a funkčně podobných druhů v dalších analýzách jsme proto použili jako mezní hodnotu 0,25 (tj, druhy s hodnotami odlišnosti < 0,25 byly považovány za funkčně společné nebo nadbytečné v rámci konkrétního biomu).
Funkční prostor a podobnost biomů
K odhadu překryvu hyperobjemů biomů jsme použili Sørensenův index podobnosti pomocí (i) celkového počtu druhů a (ii) seznamu druhů považovaných za funkčně společné a geograficky rozšířené pro každý biom. Podobnost prostoru funkčních znaků byla vypočtena jako párový frakční překryv hyperobjemů mezi biomy. Frakční překryv byl vypočten vydělením dvojnásobku objemu průsečíku dvou hyperobjemů objemem jejich spojnic. Všechny hyperobjemy byly odhadnuty pomocí balíčku R “hypervolume” (Blonder et al., 2014, 2018) s použitím Gaussovy metody KDE s výchozím Silvermanovým odhadem šířky pásma.
Výsledky
Na základě průsečíku modelovaných areálů druhů jsme zjistili značné překrývání druhového složení rostlin napříč biomy (viz obrázek 1 a doplňková tabulka 1). Toto taxonomické překrývání je větší v rámci tropických a mírných biomů, přičemž mezi těmito dvěma klimatickými pásmy je sdíleno relativně málo druhů. Zajímavé je, že xerické lesy sdílejí druhy jak s tropickými, tak s mírnými biomy. Navzdory vysokému podílu druhů charakteristických pro tropický vlhký les (83 %) sdílí tento biom velké množství druhů i s dalšími tropickými biomy, jako jsou suché lesy, savany, xerické lesy a tropické trávníky. Vysoké taxonomické překrývání biomů mírného pásma, jako jsou temperátní trávníky, středomořské lesy, tajga a tundra, naznačuje možnost nízké funkční odlišnosti, přičemž některé biomy mírného pásma představují chudší podskupinu druhově bohatších a funkčně rozmanitějších tropických biomů.
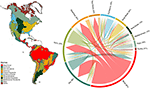
Obrázek 1. Biomy mírného pásma. Překrývání rostlinných druhů mezi biomy Nového světa. Procentuální hodnoty vyjadřují podíl druhů vyskytujících se v daném biomu, které mají největší část svého geografického areálu v tomto biomu, N = 84 413. Základna každé rozvětvené šipky je umístěna tak, aby ukazovala biom, který zahrnuje největší podíl areálu druhu, zatímco šířka představuje počet druhů sdílených s biomy na koncích šipky. Podkladová matice je uvedena v doplňkové tabulce 1.
Při zastoupení méně než 10 % mapovaných druhů byly údaje o znacích pro naše druhové soubory v měřítku biomu poměrně řídké (obr. 2A). Dostupná data o znacích se navíc značně lišila mezi jednotlivými znaky, rostlinnými klany a biomy. Z fylogenetického hlediska byly zvláště slabě zastoupeny znaky P listů a hustota dřeva, u nichž chyběla jakákoli dostupná data u celých kladů. Výsledkem bylo, že z přibližně 2/3 hodnot znaků pro našich 7842 druhů, které musely být imputovány, musely být mnohé přiřazeny při absenci jakýchkoli blízce příbuzných taxonů. Z geografického hlediska jsou SLA, výška a hmotnost semen v tropech a mírném pásmu Jižní Ameriky podhodnoceny, zatímco N v listech a hustota dřeva jsou v jednotlivých regionech podhodnoceny rovnoměrněji. Listový P je nedostatečně vzorkován ve všech biomech. Hyperobjem znaků vytvořený z druhových souborů náhodně vybraných 20 % buněk pro každý biom vykazuje rozdíly především mezi tropickými a mírnými/studenými biomy (obr. 2B). Hyperobjem znaků má tendenci být větší a variabilnější v tropických biomech, s největší variabilitou ve vlhkých a suchých tropických lesích. Mezi biomy mírného/studeného pásma mají největší hyperobjem znaků jehličnaté a smíšené lesy mírného pásma. Zajímavé je, že xerické keřové porosty vykazují rozložení hyperobjemů podobnější tropickým biomům než biomům mírného pásma.
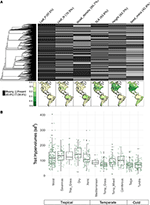
Obrázek 2. (A) Podíl druhů v databázi BIEN 3.0 se známými (šedá) nebo chybějícími (černá) hodnotami znaků. Fylogeneze vlevo odpovídá stromu ALLBM semenných rostlin od Smitha a Browna (2018) (tj. taxony GenBank a Open Tree of life s páteří poskytnutou Magallónem et al., 2015). Mapy představují podíl druhů s informacemi o znacích vzhledem k celkovému počtu druhů v databázi BIEN 2.0. (B) Rozložení hyperobjemů znaků 20 % náhodně vybraných buněk 100 × 100 km v každém biomu. Hyperobjem je uveden v jednotkách směrodatných odchylek na mocninu počtu použitých znaků.
Vztah mezi funkční odlišností a geografickým rozšířením je mezi biomy pozoruhodně podobný (obr. 3). V každém biomu je naprostá většina druhů jak geograficky omezená, tak funkčně podobná. Zároveň platí, že funkčně nejodlišnější druhy v rámci každého biomu byly zpravidla také geograficky omezené a geograficky nejrozšířenější druhy byly téměř vždy funkčně podobné.
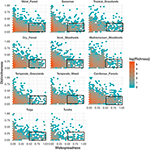
Obrázek 3. Vzorce funkční odlišnosti napříč biomy. Odlišnost představuje, jak jsou od sebe druhy v rámci daného biomu funkčně vzdáleny (tj. průměrná párová fenotypová vzdálenost ohniskového druhu od všech ostatních). Čím je hodnota větší, tím je druh vzdálenější od centroidu funkčního prostoru daného biomu. Rozšířenost měří, jak je druh v rámci biomu geograficky běžný. Hodnota 0 znamená, že se druh vyskytuje v jediné buňce biomu. Hodnoty binů představují počet druhů na logaritmické stupnici. Obdélníky představují druhy, které jsou považovány za funkčně podobné a rozšířené v každém biomu s použitím mezních hodnot odlišnosti 0,25 a rozšířenosti 0,5.
Zjistili jsme značné rozdíly v poměrném rozložení růstových forem v rámci jednotlivých biomů i mezi nimi. Dřeviny představují z hlediska počtu druhů dominantní růstové formy v tropických biomech, zatímco v mírném prostředí dominují bylinné druhy (obr. 4). Pokud jsme vzali v úvahu pouze druhy, které jsou v jednotlivých biomech rozšířené a funkčně běžné, rozložení růstových forem v jednotlivých biomech se změnilo, zejména v poměru dřevin, bylin a trav. Například v tropických biomech se podíl stromů snížil v každém biomu s výjimkou vlhkého lesa. V temperátních biomech se podíl trav zvýšil, zejména směrem k polárním oblastem. Zajímavé je, že rozložení růstových forem v xerických lesích se více podobá rozložení v temperátních biomech, pokud se berou v úvahu pouze široce rozšířené a funkčně běžné druhy.
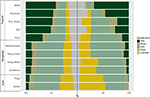
Obrázek 4: Rozložení růstových forem v xerických lesích. Rozložení růstových forem v jednotlivých biomech (vlevo) při použití celkového počtu druhů a (vpravo) při použití pouze těch druhů, které jsou funkčně podobné a rozšířené v rámci daného biomu (viz obrázek 3).
Párová srovnání druhového složení mezi biomy ukazují tři hlavní shluky reprezentující tropickou, mírnou a polární klimatickou zónu (obrázek 5A), což odráží vysoké taxonomické překrývání v rámci těchto oblastí, nikoli však mezi nimi (obrázek 1). Párová srovnání hyperobjemů znaků mezi biomy ukazují méně jasné shlukování klimatických zón (obrázek 5B). V tomto případě se funkční prostor xerických lesů výrazně překrývá s biomy mírného pásma. Tato analýza rovněž odhaluje překrývání funkčního prostoru tajgy s travnatými porosty mírného pásma a smíšenými lesy. Párové srovnání hyperobjemů znaků mezi biomy s použitím pouze těch druhů, které jsou považovány za funkčně běžné a rozšířené, ukazuje menší překrývání v prostoru znaků mezi klimatickými zónami a uvnitř nich (obr. 5C). Nicméně i když jsou nyní xerické lesy seskupeny s ostatními tropickými biomy, vykazují tyto biotopy spolu s tropickými travními porosty velký překryv ve funkčním prostoru s biomy mírného pásma, jako jsou středomořské lesy a travní porosty mírného pásma.
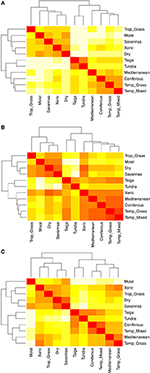
Obrázek 5. (A) Párová rozdílnost druhového složení mezi biomy. (B) Párová nepodobnost v hyperobjemnosti znaků (1-Sørensenova podobnost) mezi biomy s využitím celkového počtu druhů. (C) Párová nepodobnost v hyperobjemech znaků (1-Sørensenova podobnost) mezi biomy s použitím pouze těch druhů, které jsou považovány za funkčně podobné a rozšířené. Čím světlejší je buňka, tím větší je nepodobnost.
Diskuse
Naše analýzy přinášejí tři důležité poznatky pro pochopení suchozemských biomů funkční optikou. Za prvé ukazujeme, že navzdory pokroku v sestavování a syntéze primárních údajů o biologické rozmanitosti přetrvávají značné nedostatky ve znalostech, které mohou omezit naši schopnost kvantifikovat funkční biologickou rozmanitost biomů v kontinentálním až globálním měřítku. Za druhé, naše analýzy dostupných údajů nicméně ukazují, že všechny biomy v Severní a Jižní Americe sdílejí pozoruhodný společný vzorec, v němž geograficky nejrozšířenější druhy v každém biomu mají tendenci sdílet společné funkční znaky, zatímco funkčně nejodlišnější druhy mají vždy omezené rozšíření. Za třetí, pokud se berou v úvahu pouze široce rozšířené a funkčně společné druhy, lze biomy z funkčního hlediska snadněji rozlišit a zdá se, že vzorce odlišnosti mezi biomy odrážejí shodu mezi klimatem a prostorem funkční niky rostlin. Celkově naše výsledky naznačují, že ačkoli je studium funkční diverzity biomů stále ve své formativní fázi, další rozvoj oboru pravděpodobně přinese poznatky propojující evoluci, biogeografii, sestavu společenstev a funkci ekosystémů.
Knowledge Shortfalls
Databáze BIEN je speciálně navržena tak, aby vyplnila mezery v našich znalostech o biodiverzitě rostlin, přesto, jak Hortal et al. (2015) poukazují, vzájemně se ovlivňující nedostatky znalostí vedou k nejistotě při kvantifikaci biodiverzity v největších měřítkách. Například nejenže méně než 10 % námi zmapovaných druhů mělo k dispozici jak údaje o znacích, tak fylogenetické informace, ale tyto chybějící údaje o znacích (~66 % hodnot znaků druhů) byly značně strukturované, a to jak fylogeneticky, tak geograficky. Raunkiaerovský nedostatek dat o funkčních znacích (sensu Hortal et al., 2015) tak interaguje s wallaceovským nedostatkem distribučních informací a darwinovským nedostatkem fylogenetických znalostí. A přestože můžeme využít fylogenetické znalosti k rozumné implikaci těchto chybějících hodnot (Swenson et al., 2017), několik aspektů výsledných vzorců, které jsou důležité pro klasifikaci biomů, zůstává nejistých. Například stávající údaje o růstových formách naznačují, že dřeviny dominují ve všech tropických biomech, zatímco poměrná diverzita bylinných růstových forem je mnohem vyšší v mírných a polárních biomech (viz také Engemann et al., 2016; Zanne et al., 2018). Odráží tento latitudinální nárůst relativní diverzity bylinných forem zkreslení při výběru vzorků a nedostatek taxonomických znalostí o tropických bylinách, nebo odráží rozdílnou evoluční a biogeografickou historii v nearktické a neotropické sféře? Dominantní růstové formy jsou jedním z klíčových rozlišovacích znaků biomů, takže systematické změny nejrozmanitějších růstových forem (ať už dominantních nebo ne) nutně ovlivní naše předpovědi o tom, jak se může měnit funkční diverzita napříč biomy a jak tyto změny ovlivní funkce a služby ekosystémů.
Jako vstupní údaje jsme použili mapy areálů druhů především proto, abychom překonali nepopiratelný problém zkreslení výběru vzorků, který je charakteristický pro soubory dat sestavené z více zdrojů (viz doplňkový obrázek 1). Mapy areálů sestavené na základě modelů rozšíření druhů však mohou představovat další prvek nejistoty v našich výsledcích. Například vzhledem ke geografickému rozlišení této studie a prostorové složitosti některých biomů mohly tyto modely nadhodnotit geografický rozsah některých druhů z buněk jednoho biomu do buněk blízkých a vzájemně se prolínajících. Použití údajů o výskytu nezměnilo obecné výsledky této studie (viz doplňkové obrázky 2-6), nicméně mírné snížení funkčního překryvu mezi biomy mohlo skutečně odhalit nadhodnocení areálů některých druhů (viz doplňkové obrázky 2, 5 a 6). Použití informací o výskytu také změnilo obecné vztahy mezi funkční odlišností a rozšířeností v našich výsledcích, a to v důsledku relativního nadvýběru mírných a tropických oblastí (viz doplňkový obrázek 4). V tomto případě je naše míra rozšířenosti omezena extrémně omezeným vzorkováním většiny druhů v tropických biomech. S tím, jak se budou zlepšovat soubory dat a vzorkování, bude pravděpodobně v budoucnu existovat prostor pro snížení těchto nejistot.
Další mezerou v datech, kterou je třeba řešit pro pochopení funkční diverzity biomů, je prestonský nedostatek informací o početnosti. Početnost je při studiu funkční diverzity obzvláště důležitá, protože pro sestavování společenstev, fungování ekosystémů a biogeochemický cyklus jsou nejdůležitější znaky nejdominantnějších druhů. Protože jsme naše společenstva založili na mapách areálů odvozených z modelů rozšíření druhů, mohli jsme se zabývat pouze geografickou složkou rozšířenosti, ale pozitivní vztah mezi lokální početností a geografickým rozsahem, tzv. vztah obsazenosti a hojnosti, naznačuje, že nejrozšířenější druhy v každém biomu mohou často patřit také k těm hojnějším (Borregaard a Rahbek, 2010; Novosolov et al., 2017). Vzhledem k tomu, že biomy jsou obecně charakterizovány dominantními růstovými formami v daném regionu, může integrace informací o početnosti vést k tomu, že biomy budou vykazovat menší funkční překryv. Nicméně i když BIEN shromáždila objemné údaje o plochách, které umožňují odhady lokální početnosti, provedení rozumného srovnání mezi různými regiony a růstovými formami brání nerovnoměrný odběr vzorků, regionální rozdíly v diverzitě gama a nesouměřitelné metody kvantifikace početnosti.
Společné vzorce v rámci biomů
Přes přetrvávající mezery v dostupných údajích naše analýzy odhalily dříve nedokumentovaný vztah společný pro všechny biomy západní polokoule: v každém biomu mají nejrozšířenější druhy také tendenci vykazovat nízkou funkční odlišnost, zatímco funkčně nejodlišnější druhy mají téměř vždy omezené rozšíření (obr. 3). Tento vztah “obsazenosti a redundance” může naznačovat, že klimatické podmínky převládající v daném biomu vybírají soubor společných charakteristik, přičemž funkčně odlišnější druhy jsou omezeny na vzácnější podskupinu stanovišť v rámci daného biomu. Celková převaha funkčně podobných druhů ve všech biomech a vztah “obsazenost-redundance” jsou v souladu s nedávnou globální analýzou funkční diverzity na úrovni společenstev, která naznačuje, že filtrování stanovišť vede ke koexistenci funkčně podobných druhů (Li et al., 2018), a také se studiemi, které ukazují, že funkční redundance zvyšuje stabilitu společenstev (např. Walker et al., 1999; Pillar et al., 2013). Navíc spolu s vysokou mírou taxonomického i funkčního překrývání mezi biomy (obr. 1, 5) skutečnost, že běžné, široce rozšířené druhy jsou si funkčně podobné, posiluje představu, že suchozemské rostliny v širokém spektru podmínek prostředí sdílejí společné vlastnosti blízko jádra spektra funkčních znaků (Wright et al., 2004; Díaz et al., 2016). Na rozdíl od Umaña et al. (2017) naše výsledky ukazují, že vzácné, geograficky omezené druhy se mohou, ale nemusí funkčně lišit od rozšířenějších a běžných druhů. Zůstává tedy otevřená otázka, zda jsou funkčně odlišnější druhy specialisty na určitá prostředí, nebo zda kombinace jejich znaků vede k demografickým či fyziologickým kompromisům, které omezují jejich geografické rozšíření.
Srovnání mezi biomy
Naše srovnání rozložení hyperobjemů znaků mezi biomy (obr. 2B) je v souladu s pozorováním, že druhově bohatší prostředí jsou také funkčně rozmanitější (Swenson et al., 2012; Lamanna et al., 2014; Li et al., 2018; Šímová et al., 2018). Naše výsledky jsou nejednoznačnější, pokud jde o hypotézu, že sezónní a extrémní klimatická prostředí důsledně omezují funkční rozmanitost druhů (de la Riva et al., 2018). Všechny tropické biomy vykazují vyšší průměrnou funkční diverzitu než všechny biomy mírného pásma a polární biomy skutečně vykazují nejmenší hyperobjem. V rámci každé skupiny však sušší nebo sezónně proměnlivější biomy nevykazují vždy menší hyperobjem (např. suché lesy, xerické lesy), jak bychom očekávali podle hypotézy tolerance Currieho et al. (2004), zatímco optimální klimatické podmínky podporují více kombinací fyziologických parametrů. Pouze z těchto výsledků nemůžeme určit, zda jsou biomy mírného pásma a polární biomy méně taxonomicky rozmanité kvůli limitům funkčního prostoru niky, nebo zda jsou jejich funkční hyperobjemy malé, protože nejsou taxonomicky rozmanité. Přístupy nulového modelování by potenciálně mohly pomoci oddělit taxonomickou a funkční diverzitu (Swenson et al., 2012; Lamanna et al., 2014; Šímová et al., 2018), ale taková analýza byla nad rámec této studie. Důležitější je, že naše výsledky posilují význam pochopení toho, jak evoluční a biogeografická historie formuje funkční diverzitu biomů (Woodward et al., 2004; Pennington et al., 2006; Donoghue a Edwards, 2014; Moncrieff et al., 2016). Rozsáhlé sdílení druhů (a vyšších linií) napříč biomy v rámci různých biogeografických oblastí, ale převážně ne mezi nimi, by mohlo sloužit jak k homogenizaci funkční diverzity v rámci oblastí, tak k poskytnutí vodítek o charakteristických rysech, které jsou různými prostředími selektovány, resp. proti nimž jsou vybírány (Douma et al., 2012; Zanne et al., 2014, 2018).
Přes značné překrývání hyperobjemů mezi všemi biomy (doplňkový obrázek 2) se zdá, že tropické, mírné a chladné biomy zaujímají odlišné oblasti funkčního prostoru (obrázky 5B,C). Hlavními znaky odlišujícími biomy se zdají být znaky související s celkovou velikostí rostlin, včetně vzrostlé výšky i hmotnosti semen (doplňkové obrázky 7-10), spíše než znaky ekonomiky listů, jak bylo pozorováno v lokálnějších analýzách založených na plochách (např. Douma et al., 2012). Výjimkou je listové P, které vykazovalo značné rozdíly mezi tropickými a mírnými/polárními biomy (doplňkové obrázky 7-10), což je vzorec, který byl pozorován v jiných analýzách založených jak na druhově specifických hodnotách, tak na měřeních celého ekosystému (Kerkhoff et al., 2005; Swenson et al., 2012). Vzhledem k tomu, že listové P bylo naším nejvzorkovanějším znakem, rozdíly mezi tropickými a mírnými/polárními oblastmi mohou podléhat zkreslení způsobenému postupem imputace. Nicméně list P vykazuje významný fylogenetický signál (Kerkhoff et al., 2006), což naznačuje, že imputace by měly být nezkreslené (Swenson et al., 2017). Leaf P je také významně vyšší u bylinných růstových forem (Kerkhoff et al., 2006), takže latitudinální posun od převážně dřevinné k bylinné diverzitě (obr. 4) může mít vliv i na tento vzorec.
Když jsme omezili analýzy hyperobjemů pouze na nejrozšířenější a funkčně nejběžnější druhy v každém biomu, jednotlivé biomy v rámci každé biogeografické oblasti se méně překrývaly v prostoru funkčních znaků (srovnej obr. 5B,C), což naznačuje, že tyto druhy mohou odrážet fenotypy lépe přizpůsobené určitým prostředím. V této souvislosti je zvláště zajímavý biom xerických lesů. Částečně díky zahrnutí ekoregionu Chaco se xerické lesy taxonomicky shlukují s tropickými biomy (obr. 5A). Pokud však vezmeme v úvahu funkční hyperobjem všech druhů, vykazují mnohem větší podobnost s biomy mírného pásma (obr. 5B). A konečně, pokud se analyzují pouze nejrozšířenější a funkčně nejběžnější druhy, xerické lesy opět vykazují zvýšenou podobnost s tropickými biomy, ale zároveň si zachovávají vysokou podobnost s mírnými travinnými a středomořskými biomy (obrázek 5C). Přechody z teplých, mezických prostředí do chladnějších, sušších a sezónních prostředí usnadňují podobné znaky, např. bylinný habitus (Douma et al., 2012; Zanne et al., 2014, 2018), a stejně jako chladnější prostředí mají i společenstva v suchých prostředích tendenci být fylogeneticky shlukovanější (Qian a Sandel, 2017). Poloha xerických zón na hranici intertropické konvergenční zóny z nich navíc činí geografický přechod mezi tropickou a mírnou oblastí. Skutečnost, že xerické lesy jsou funkčně i biogeograficky na pomezí tropické a mírné říše, dále posiluje myšlenku, že pro lepší pochopení funkční diverzity biomů musíme brát v úvahu jejich biogeografickou a fylogenetickou historii (Pennington et al., 2006; Donoghue a Edwards, 2014; Moncrieff et al., 2016).
Závěry
Každá klasifikace suchozemských biomů vnucuje spojitým gradientům klimatu a rozšíření druhů malý počet diskrétních kategorií, a představuje tak hrubé zjednodušení složité ekologické krajiny (Moncrieff et al., 2016, ale viz Silva de Miranda et al., 2018). I přes potenciál přílišného zjednodušení jsou však biomy užitečnými konstrukty pro uspořádání a pochopení biodiverzity a fungování hlavních suchozemských ekosystémů na Zemi a přístupy založené na znacích mají vysoký potenciál pomoci dynamicky modelovat globální rozšíření vegetace.
V této studii jsme ukázali, že velká část taxonomické rozmanitosti všech biomů představuje druhy, které jsou úzce rozšířené a zároveň funkčně podobné. Dále, v rámci biomů mají nejvíce funkčně odlišné druhy v každém biomu také tendenci být geograficky vzácné, zatímco široce rozšířené druhy shodně vykazují nízkou funkční odlišnost. Navzdory rozsáhlému taxonomickému a funkčnímu překrývání mezi biomy se tyto shlukují do rozlišitelných, biogeograficky a klimaticky odlišných jednotek, zejména pokud je funkční shlukování založeno na nejrozšířenějších a funkčně nejběžnějších druzích v každém biomu. Pokrok ve funkčním chápání biomů však bude vyžadovat nejen lepší charakterizaci variability znaků v rámci vegetačních typů a mezi nimi, ale také informace o biogeografické a fylogenetické historii druhových společenstev a relativní početnosti druhů v rámci biomů.
Dostupnost dat
Soubory dat o znacích a mapy rozšíření druhů analyzované pro tuto studii lze stáhnout prostřednictvím balíčku R BIEN (Maitner et al., 2018). Více informací o databázi BIEN viz http://bien.nceas.ucsb.edu/bien/.
Příspěvky autorů
SE-L, AK, BJE, DMN a CV navrhli studii; SE-L a AK analyzovali data; SE-L a AK napsali první verzi rukopisu; CV, BB, NK, BSM, BM, RKP, BS, SS, J-CS a SKW přispěli ke srovnání a vytvoření databáze znaků a map rozšíření. Všichni autoři se podíleli na vývoji a psaní rukopisu.
Financování
SE-L, DMN a AK byli podpořeni grantem na společný výzkum od US National Science Foundation (DEB-1556651). BJE byl podpořen grantem National Science Foundation DEB-1457812 a Macrosystems-1065861. CV byl podpořen Evropskou výzkumnou radou (grant ERC-StG-2014-639706-CONSTRAINTS) a Francouzskou nadací pro výzkum biodiverzity (FRB; www.fondationbiodiversite.fr) v rámci projektu CESAB Causes and consequences of functional rarity from local to global scales. J-CS považuje tuto práci za příspěvek ke svému projektu VILLUM Investigator Biodiversity Dynamics in a Changing World financovanému VILLUM FONDEN (grant 16549). SKW byl podpořen Strategic Science Investment Fund of the New Zealand Ministry of Business, Innovation and Employment’s Science and Innovation Group.
Prohlášení o střetu zájmů
Autoři prohlašují, že výzkum byl prováděn bez jakýchkoli komerčních nebo finančních vztahů, které by mohly být chápány jako potenciální střet zájmů.
Poděkování
Děkujeme Danu Griffithovi a dvěma recenzentům za jejich konstruktivní kritiku, která výrazně zlepšila tento rukopis. Děkujeme také všem přispěvatelům dat BIEN (úplný seznam naleznete v části http://bien.nceas.ucsb.edu/bien/data-contributors/).
Přídavné materiály
Přídavné materiály k tomuto článku naleznete online na adrese: https://www.frontiersin.org/articles/10.3389/fevo.2018.00219/full#supplementary-material
Banda, R.K., Delgado-Salinas, A., Dexter, K. G., Linares-Palomino, R., Oliveira-Filho, A., Prado, D., et al. (2016). Modely diverzity rostlin v neotropických suchých lesích a jejich důsledky pro ochranu přírody. Science 353, 1383-1387. doi: 10.1126/science.aaf5080
PubMed Abstract | CrossRef Full Text | Google Scholar
Blonder, B., Lamanna, C., Violle, C., and Enquist, B. J. (2014). N-rozměrný hyperobjem. Glob. Ecol. Biogeogr. 23, 595-609. doi: 10.1111/geb.12146
CrossRef Full Text | Google Scholar
Blonder, B., Morrow, C. B., Maitner, B., Harris, D. J., Lamanna, C., Violle, C. a další (2018). Nové přístupy k vymezování n-rozměrných hyperobjemů. Methods Ecol. Evol. 9, 305-319. doi: 10.1111/2041-210X.12865
CrossRef Full Text | Google Scholar
Bonan, G. B., Oleson, K. W., Fisher, R. A., Lasslop, G., and Reichstein, M. (2012). Sladění fyziologických vlastností listů a údajů o tocích v korunách stromů: použití databází TRY a FLUXNET v modelu Společenství země verze 4. J. Geophys. Res. Biogeosci. 117:G02026. doi: 10.1029/2011JG001913
CrossRef Full Text | Google Scholar
Borregaard, M. K., and Rahbek, C. (2010). Kauzalita vztahu mezi geografickým rozšířením a početností druhů. Q. Rev. Biol. 85, 3-25. doi: 10.1086/650265
PubMed Abstract | CrossRef Full Text | Google Scholar
Bruggeman, J., Heringa, J., and Brandt, B. W. (2009). PhyloPars: odhad chybějících hodnot parametrů pomocí fylogeneze. Nucleic Acids Res. 37, W179-W184. doi: 10.1093/nar/gkp370
PubMed Abstract | CrossRef Full Text | Google Scholar
Cadotte, M. W., Carscadden, K., and Mirotchnick, N. (2011). Beyond species: functional diversity and the maintenance of ecological processes and services (Za hranice druhů: funkční rozmanitost a zachování ekologických procesů a služeb). J. Appl. Ecol. 48, 1079-1087. doi: 10.1111/j.1365-2664.2011.02048.x
CrossRef Full Text | Google Scholar
Cornwell, W. K., Cornelissen, J. H., Amatangelo, K., Dorrepaal, E., Eviner, V. T., Godoy, O., et al. (2008). Plant species traits are the predominant control on litter decomposition rates within biomes worldwide (Vlastnosti rostlinných druhů jsou dominantním faktorem ovlivňujícím rychlost rozkladu steliva v rámci světových biomů). Ecol. Lett. 11, 1065-1071. doi: 10.1111/j.1461-0248.2008.01219.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Craine, J. M. (2009). Resource Strategies of Wild Plants (Zdrojové strategie planě rostoucích rostlin). New Jersey, NJ: Princeton University Press. doi: 10.1515/9781400830640
CrossRef Full Text | Google Scholar
Currie, D. J., Mittelbach, G. G., Cornell, H. V., Field, R., Guégan, J. F., Hawkins, B. A., et al. (2004). Predictions and tests of climate-based hypotheses of broad-scale variation in taxonomic richness [Předpovědi a testování hypotéz o variabilitě taxonomického bohatství na základě klimatu]. Ecol. Lett. 12, 1121-1134. doi: 10.1111/j.1461-0248.2004.00671.x
CrossRef Full Text | Google Scholar
de la Riva, E. G., Violle, C., Pérez-Ramos, I. M., Marañón, T., Navarro-Fernández, C. M., Olmo, M., et al. (2018). A multidimensional functional trait approach reveals the imprint of environmental stress in Mediterranean woody Communities [Vícerozměrný přístup k funkčním znakům odhaluje otisk environmentálního stresu u středomořských dřevin]. Ecosystems 21, 248-262. doi: 10.1007/s10021-017-0147-7
CrossRef Full Text | Google Scholar
Díaz, S., Kattge, J., Cornelissen, J. H., Wright, I. J., Lavorel, S., Dray, S., et al. (2016). Globální spektrum tvarů a funkcí rostlin. Nature 529, 167-171. doi: 10.1038/nature16489
PubMed Abstract | CrossRef Full Text | Google Scholar
Díaz, S., Lavorel, S., de Bello, F., Quétier, F., Grigulis, K., and Robson, T. M. (2007). Zahrnutí vlivu funkční rozmanitosti rostlin do hodnocení ekosystémových služeb. Proc. Natl. Acad. Sci. U.S.A 104, 20684-20689. doi: 10.1073/pnas.0704716104
PubMed Abstract | CrossRef Full Text | Google Scholar
Donoghue, M. J., and Edwards, E. J. (2014). Biome shifts and niche evolution in plants (Změny biomů a evoluce nik u rostlin). Annu. Rev. Ecol. Evol. Syst. 45, 547-572. doi: 10.1146/annurev-ecolsys-120213-091905
CrossRef Full Text | Google Scholar
Douma, J. C., Aerts, R., Witte, J. P. M., Bekker, R. M., Kunzmann, D., Metselaar, K., et al. (2012). Kombinace funkčně odlišných znaků rostlin poskytuje prostředek ke kvantitativní předpovědi široké škály druhových společenstev v SZ Evropě. Ecography 35, 364-373. doi: 10.1111/j.1600-0587.2011.07068.x
CrossRef Full Text | Google Scholar
Engemann, K., Sandel, B., Enquist, B. J., Jørgensen, P. M., Kraft, N., Marcuse-Kubitza, A., et al. (2016). Patterns and drivers of plant functional group dominance across the Western Hemisphere: a macroecological re-assessment based on a massive botanical dataaset (Vzorce a faktory dominance funkčních skupin rostlin na západní polokouli: makroekologické přehodnocení na základě rozsáhlého souboru botanických dat). Bot. J. Linn. Soc. 180, 141-160. doi: 10.1111/boj.12362
CrossRef Full Text | Google Scholar
Enquist, B. J., Condit, R., Peet, R., Thiers, B., and Schildhauer, M. (2016). The botanical information and ecology network (BIEN): cyberinfrastructure for an integrated botanical information network to investigate the ecological impacts of global climate change on plant biodiversity. PeerJ Prepr. 4:e2615v2. doi: 10.7287/peerj.preprints.2615v2
CrossRef Full Text | Google Scholar
Garnier, E., Cortez, J., Billès, G., Navas, M. L., Roumet, C., Debussche, M., et al. (2004). Funkční markery rostlin zachycují vlastnosti ekosystémů během sekundární sukcese. Ecology 85, 2630-2637. doi: 10.1890/03-0799
CrossRef Full Text | Google Scholar
Goldsmith, G. R., Morueta-Holme, N., Sandel, B., Fitz, E. D., Fitz, S. D., Boyle, B., et al. (2016). Plant-O-Matic: dynamický a mobilní průvodce všemi rostlinami Ameriky. Methods Ecol. Evol. 7, 960-965. doi: 10.1111/2041-210X.12548
CrossRef Full Text | Google Scholar
Goolsby, E. W., Bruggeman, J., and Ané, C. (2017). Rphylopars: rychlé vícerozměrné fylogenetické srovnávací metody pro chybějící data a vnitrodruhovou variabilitu. Methods Ecol. Evol. 8, 22-27. doi: 10.1111/2041-210X.12612
CrossRef Full Text | Google Scholar
Grenié, M., Denelle, P., Tucker, C. M., Munoz, F., and Violle, C. (2017). funrar: an R package to characterize functional rarity. Divers. Distrib. 23, 1365-1371. doi: 10.1111/ddi.12629
CrossRef Full Text | Google Scholar
Hortal, J., de Bello, F., Diniz-Filho, J. A. F., Lewinsohn, T. M., Lobo, J. M., and Ladle, R. J. (2015). Sedm nedostatků, které sužují rozsáhlé znalosti o biologické rozmanitosti. Annu. Rev. Ecol. Evol. Syst. 46, 523-549. doi: 10.1146/annurev-ecolsys-112414-054400
CrossRef Full Text | Google Scholar
Kerkhoff, A. J., Enquist, B. J., Elser, J. J., and Fagan, W. F. (2005). Rostlinná alometrie, stechiometrie a závislost primární produktivity na teplotě. Glob. Ecol. Biogeogr. 14, 585-598. doi: 10.1111/j.1466-822X.2005.00187.x
CrossRef Full Text | Google Scholar
Kerkhoff, A. J., Fagan, W. F., Elser, J. J., and Enquist, B. J. (2006). Fylogenetická variabilita a variabilita růstových forem ve škálování dusíku a fosforu u semenných rostlin. Am. Nat. 168, E103-E122. doi: 10.1086/507879
PubMed Abstract | CrossRef Full Text | Google Scholar
Kraft, N. J., Valencia, R., and Ackerly, D. D. (2008). Functional traits and niche-based tree community assembly in an Amazonian forest (Funkční vlastnosti a sestavování společenstev stromů v amazonském pralese na základě nik). Science 322, 580-582. doi: 10.1126/science.1160662
PubMed Abstract | CrossRef Full Text | Google Scholar
Lamanna, C., Blonder, B., Violle, C., Kraft, N. J., Sandel, B., et al. (2014). Prostor funkčních znaků a gradient diverzity v zeměpisné šířce. Proc. Natl. Acad. Sci. U.S.A. 111, 13745-13750. doi: 10.1073/pnas.1317722111
PubMed Abstract | CrossRef Full Text | Google Scholar
Lavorel, S., and Garnier, E. (2002). Předpovídání změn ve složení společenstev a fungování ekosystémů na základě znaků rostlin: návrat ke svatému grálu. Funct. Ecol. 16, 545-556. doi: 10.1046/j.1365-2435.2002.00664.x
CrossRef Full Text | Google Scholar
Lavorel, S., Grigulis, K., Lamarque, P., Colace, M.-P., Garden, D., Girel, J., et al. (2011). Využití funkčních znaků rostlin k pochopení rozložení vícenásobných ekosystémových služeb v krajině. J. Ecol. 99, 135-147. doi: 10.1111/j.1365-2745.2010.01753.x
CrossRef Full Text | Google Scholar
Li, Y., Shipley, B., Price, J. N., Dantas, V. de L., Tamme, R., Westoby, M., et al. (2018). Habitat filtering determines the functional niche occupancy of plant communities worldwide [Filtrování stanovišť určuje obsazení funkčních nik rostlinných společenstev po celém světě]. J. Ecol. 106, 1001-1009. doi: 10.1111/1365-2745.12802
CrossRef Full Text | Google Scholar
Magallón, S., Gómez-Acevedo, S., Sánchez-Reyes, L. L., and Hernández-Hernández, T. (2015). Metakalibrovaný časový strom dokumentuje raný vzestup fylogenetické diverzity kvetoucích rostlin. New Phytol. 207, 437-453. doi: 10.1111/nph.13264
PubMed Abstract | CrossRef Full Text | Google Scholar
Maitner, B. S., Boyle, B., Casler, N., Condit, R., Donoghue, J., Durán, S. M., et al. (2018). The BIEN R package: a tool to access the botanical information and ecology network (BIEN) database (Balíček BIEN v jazyce R: nástroj pro přístup k databázi botanických informací a ekologie (BIEN)). Methods Ecol. Evol. 9, 373-379. doi: 10.1111/2041-210X.12861
CrossRef Full Text | Google Scholar
Moncrieff, G. R., Bond, W. J., and Higgins, S. I. (2016). Revize konceptu biomů pro pochopení a předvídání dopadů globálních změn. J. Biogeogr. 43, 863-873. doi: 10.1111/jbi.12701
CrossRef Full Text | Google Scholar
Novosolov, M., Rodda, G. H., North, A. C., Butchart, S. H. M., Tallowin, O. J. S., Gainsbury, A. M., et al. (2017). Population density-range size relationship revisited. Glob. Ecol. Biogeogr. 26, 1088-1097. doi: 10.1111/geb.12617
CrossRef Full Text | Google Scholar
Olson, D. M., Dinerstein, E., Wikramanayake, E. D., Burgess, N. D., Powell, G. V. N., Underwood, E. C., et al. (2001). Terestrické ekoregiony světa: nová mapa života na Zemi: nová globální mapa terestrických ekoregionů poskytuje inovativní nástroj pro zachování biologické rozmanitosti. Bioscience 51, 933-938.doi: 10.1641/0006-3568(2001)0512.0.CO;2
CrossRef Full Text | Google Scholar
Pennington, R. T., Prado, D. E., and Pendry, C. A. (2000). Neotropické sezónně suché lesy a kvartérní změny vegetace. J. Biogeogr. 27, 261-273. doi: 10.1046/j.1365-2699.2000.00397.x
CrossRef Full Text | Google Scholar
Pennington, R. T., Richardson, J. E., and Lavin, M. (2006). Pohled na historickou stavbu druhově bohatých biomů na základě datovaných fylogenezí rostlin, neutrální ekologické teorie a fylogenetické struktury společenstev. New Phytol. 172, 605-616. doi: 10.1111/j.1469-8137.2006.01902.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Penone, C., Davidson, A. D., Shoemaker, K. T., Di Marco, M., Rondinini, C., Brooks, T. M., et al. (2014). Imputace chybějících údajů v souborech údajů o životních vlastnostech: který přístup funguje nejlépe? Methods Ecol. Evol. 5, 961-970. doi: 10.1111/2041-210X.12232
CrossRef Full Text | Google Scholar
Pillar, V. D., Blanco, C. C., Müller, S. C., Sosinski, E. E., Joner, F., and Duarte, L. D. S. (2013). Funkční redundance a stabilita v rostlinných společenstvech. J. Veg. Sci. 24, 963-974. doi: 10.1111/jvs.12047
CrossRef Full Text | Google Scholar
Prado, D. E., and Gibbs, P. E. (1993). Patterns of species distributions in the dry seasonal forests of South America [Vzorce rozšíření druhů v suchých sezónních lesích Jižní Ameriky]. Ann. Mo. Bot. Gard. 80, 902-927. doi: 10.2307/2399937
CrossRef Full Text | Google Scholar
Qian, H., and Sandel, B. (2017). Fylogenetická struktura regionálních společenstev nahosemenných rostlin napříč šířkovými a klimatickými gradienty v Severní Americe. Glob. Ecol. Biogeogr. 26, 1258-1269. doi: 10.1111/geb.12634
CrossRef Full Text | Google Scholar
Reich, P. B., Wright, I. J., Cavender-Bares, J., Craine, J. M., Oleksyn, J., Westoby, M., et al. (2003). The evolution of plant functional variation: traits, spectra, and strategies (Evoluce funkční variability rostlin: znaky, spektra a strategie). Int. J. Plant Sci. 164, S143-S164. doi: 10.1086/374368
CrossRef Full Text | Google Scholar
Silva de Miranda, P. L., Oliveira-Filho, A., Pennington, R. T., Neves, D. M., Baker, T. R., and Dexter, K. G. (2018). Using tree species inventories to map biomes and assess their climatic overlaps in lowland tropical South America [Využití inventarizace dřevin k mapování biomů a hodnocení jejich klimatických překryvů v nížinných oblastech Jižní Ameriky]. Glob. Ecol. Biogeogr. 27, 899-912. doi: 10.1111/geb.12749
CrossRef Full Text | Google Scholar
Šímová, I., Violle, C., Kraft, N. J. B., Storch, D., Svenning, J.-C., Boyle, B., et al. (2015). Shifts in trait means and variances in North American tree assemblages: species richness patterns are loosely related to the functional space. Ecography 38, 649-658. doi: 10.1111/ecog.00867
CrossRef Full Text | Google Scholar
Šímová, I., Violle, C., Svenning, J.-C., Kattge, J., Engemann, K., Sandel, B., et al. (2018). Prostorové vzorce a klimatické vztahy hlavních rostlinných znaků v Novém světě se liší mezi dřevinami a bylinami. J. Biogeogr. 45, 895-916. doi: 10.1111/jbi.13171
CrossRef Full Text | Google Scholar
Smith, S. A., and Beaulieu, J. M. (2009). Life history influences rates of climatic niche evolution in flowering plants (Životní historie ovlivňuje rychlost vývoje klimatických nik u kvetoucích rostlin). Proc. Biol. Sci. 276, 4345-4352. doi: 10.1098/rspb.2009.1176
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, S. A., and Brown, J. W. (2018). Constructing a broadly inclusive seed plant phylogeny (Konstrukce široce zahrnující fylogeneze semenných rostlin). Am. J. Bot. 105, 302-314. doi: 10.1002/ajb2.1019
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, S. A., and Donoghue, M. J. (2008). Rychlost molekulární evoluce souvisí s historií života u kvetoucích rostlin. Science 322, 86-89. doi: 10.1126/science.1163197
PubMed Abstract | CrossRef Full Text | Google Scholar
Swenson, N. G. (2014). Phylogenetic imputation of plant functional trait databases. ecography 37, 105-110. doi: 10.1111/j.1600-0587.2013.00528.x
CrossRef Full Text
Swenson, N. G., Enquist, B. J., Pither, J., Kerkhoff, A. J., Boyle, B., Weiser, M. D., et al. (2012). Biogeografie a filtrace funkční diverzity dřevin v Severní a Jižní Americe. Glob. Ecol. Biogeogr. 21, 798-808. doi: 10.1111/j.1466-8238.2011.00727.x
CrossRef Full Text | Google Scholar
Swenson, N. G., Weiser, M. D., Mao, L., Araújo, M. B., Diniz-Filho, J. A. F., Kollmann, J., et al. (2017). Fylogeneze a predikce funkční diverzity stromů v nových kontinentálních podmínkách. Glob. Ecol. Biogeogr. 26, 553-562. doi: 10.1111/geb.12559
CrossRef Full Text | Google Scholar
Umaña, M. N., Mi, X., Cao, M., Enquist, B. J., Hao, Z., Howe, R., et al. (2017). Úloha funkční jedinečnosti a prostorové agregace při vysvětlování vzácnosti stromů. Glob. Ecol. Biogeogr. 26, 777-786. doi: 10.1111/geb.12583
CrossRef Full Text | Google Scholar
van Bodegom, P. M., Douma, J. C., and Verheijen, L. M. (2014). Přístup k modelování globálního rozšíření vegetace plně založený na vlastnostech. Proc. Natl. Acad. Sci. U.S.A. 111, 13733-13738. doi: 10.1073/pnas.1304551110
PubMed Abstract | CrossRef Full Text | Google Scholar
Violle, C., Reich, P. B., Pacala, S. W., Enquist, B. J., and Kattge, J. (2014). Vznik a příslib funkční biogeografie. Proc. Natl. Acad. Sci. U.S.A. 111, 13690-13696. doi: 10.1073/pnas.1415442111
PubMed Abstract | CrossRef Full Text | Google Scholar
Violle, C., Thuiller, W., Mouquet, N., Munoz, F., Kraft, N. J. B., Cadotte, M. W., et al. (2017). Functional rarity: the ecology of outliers (Funkční vzácnost: ekologie odlehlých druhů). Trends Ecol. Evol. 32, 356-367. doi: 10.1016/j.tree.2017.02.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Walker, B., Kinzig, A., and Langridge, J. (1999). Rozmanitost atributů rostlin, odolnost a funkce ekosystému: povaha a význam dominantních a minoritních druhů. Ecosystems 2, 95-113. doi: 10.1007/s100219900062
CrossRef Full Text | Google Scholar
Westoby, M. (1998). Schéma strategie ekologie rostlin podle výšky listů (LHS). Plant Soil 199, 213-227. doi: 10.1023/A:1004327224729
CrossRef Full Text | Google Scholar
Woodward, F. I., Lomas, M. R., and Kelly, C. K. (2004). Globální klima a rozšíření rostlinných biomů. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 359, 1465-1476. doi: 10.1098/rstb.2004.1525
PubMed Abstract | CrossRef Full Text | Google Scholar
Wright, I. J., Reich, P. B., Westoby, M., Ackerly, D. D., Baruch, Z., Bongers, F., et al. (2004). Celosvětové spektrum listové ekonomiky. Nature 428, 821-827. doi: 10.1038/nature02403
PubMed Abstract | CrossRef Full Text | Google Scholar
Xia, J., Niu, S., Ciais, P., Janssens, I. A., Chen, J., Ammann, C., et al. (2015). Společná kontrola suchozemské hrubé primární produktivity fenologií a fyziologií rostlin. Proc. Natl. Acad. Sci. U.S.A. 112, 2788-2793. doi: 10.1073/pnas.1413090112
PubMed Abstract | CrossRef Full Text | Google Scholar
Zanne, A. E., Pearse, W. D., Cornwell, W. K., McGlinn, D. J., Wright, I. J., and Uyeda, J. C. (2018). Functional biogeography of angiosperms: life at the extremes [Funkční biogeografie nahosemenných rostlin: život v extrémech]. New Phytol. 218, 1697-1709. doi: 10.1111/nph.15114
PubMed Abstract | CrossRef Full Text | Google Scholar
Zanne, A. E., Tank, D. C., Cornwell, W. K., Eastman, J. M., Smith, S. A., FitzJohn, R. G., et al. (2014). Tři klíče k radiaci nahosemenných rostlin do mrazových prostředí. Nature 506, 89-92. doi: 10.1038/nature12872
PubMed Abstract | CrossRef Full Text | Google Scholar
.