Hodnocení | Biopsychologie | Srovnávací | Kognitivní | Vývojová | Jazyk | Individuální rozdíly | Osobnost | Filozofie | Sociální |
Metody | Statistika | Klinická | Vzdělávací | Průmyslová | Odborné předměty | Světová psychologie |
Biologická:Neuroanatomie – Neurochemie – Neuroendokrinologie -Neurologie – Psychoneuroimunologie – Fyziologická psychologie – Psychofarmakologie(Rejstřík, Osnova)
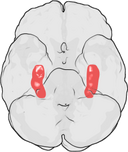
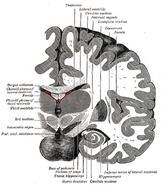
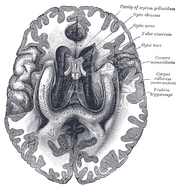
Umístění hipokampu v lidském mozku.
Pro stejnojmenný časopis viz Hippocampus (časopis)
Hipokampus je část předního mozku, která se nachází v mediálním spánkovém laloku. Patří do limbického systému a hraje hlavní roli v dlouhodobé paměti a prostorové navigaci. Lidé a další savci mají dva hipokampy, na každé straně mozku jeden. U hlodavců, kde byl studován nejrozsáhleji, má hipokampus tvar podobný banánu. U člověka má zakřivený a stočený tvar, který prvním anatomům připomínal mořského koníka. Název je ve skutečnosti odvozen od řeckého slova pro mořského koníka (řecky: hippos = kůň, campos = moře).
Při Alzheimerově chorobě je hipokampus jednou z prvních oblastí mozku, která je poškozena; mezi prvními příznaky se objevují problémy s pamětí a dezorientace. Poškození hipokampu může být také důsledkem nedostatku kyslíku (anoxie), encefalitidy nebo epilepsie mesiálního temporálního laloku. U lidí s rozsáhlým poškozením hipokampu se může vyskytnout amnézie, tj. neschopnost vytvářet nebo uchovávat nové vzpomínky.
Funkce hipokampu
Snad nejstarší myšlenkou bylo, že hipokampus se podílí na čichu: zdá se, že to bylo naznačeno především jeho umístěním v mozku, vedle čichové kůry. Zájem o čichové reakce hipokampu přetrvává, ale téměř nikdo už nevěří, že primární funkcí hipokampu je čich.
V průběhu let v literatuře převládly tři hlavní představy o funkci hipokampu: inhibice, paměť a prostor. Teorie behaviorální inhibice (karikovaná O’Keefem a Nadelem jako “šlápněte na brzdu!”) byla až do 60. let 20. století velmi populární. Velkou část své síly odvozovala ze dvou pozorování: za prvé, zvířata s poškozením hipokampu bývají hyperaktivní; za druhé, zvířata s poškozením hipokampu mají často potíže naučit se inhibovat reakce, které se předtím naučila. Jeffrey Gray tuto myšlenkovou linii rozvinul do plnohodnotné teorie o roli hipokampu v úzkosti. Teorie inhibice však není v současné době příliš populární.
Druhá důležitá myšlenková linie souvisí s hipokampem a pamětí. Ačkoli měla své předchůdce, tato myšlenka čerpala svou hlavní sílu z velmi známé zprávy Scovilla a Milnera o výsledcích chirurgické destrukce hipokampu (ve snaze zmírnit epileptické záchvaty) u pacienta známého jako H.M. Nečekaným výsledkem byla těžká amnézie: H.M. si nebyl schopen vědomě vzpomenout na události, které se staly po operaci nebo několik let před ní. Tento případ vyvolal tak obrovský zájem, že H. M. je dnes považován za nejintenzivněji studovaný lékařský případ v historii. V následujících letech byli studováni i další pacienti s podobným stupněm poškození hipokampu a amnézií (způsobenou nehodou nebo nemocí) a doslova tisíce experimentů studovaly fyziologii nervové plasticity v hipokampu. V současné době panuje téměř všeobecná shoda na tom, že hipokampus hraje v paměti určitou důležitou roli; o přesné povaze této role se však stále vedou rozsáhlé diskuse.
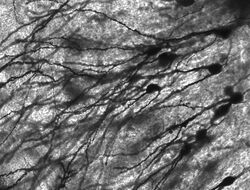
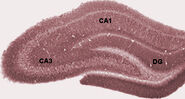
Golgiho zbarvené neurony v hipokampu hlodavců.
Role v paměti
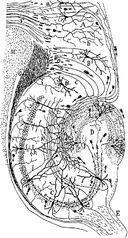
Kresba nervového obvodu hipokampu hlodavců. S. Ramón y Cajal, 1911.
Psychologové a neurovědci se přou o přesnou roli hipokampu, ale obecně se shodují, že má zásadní úlohu při vytváření nových vzpomínek na osobně prožité události (epizodická nebo autobiografická paměť). Někteří badatelé dávají přednost tomu, aby byl hipokampus považován za součást většího paměťového systému mediálního temporálního laloku, který je zodpovědný za obecnou deklarativní paměť (vzpomínky, které lze explicitně verbalizovat – mezi ně by kromě epizodické paměti patřila například paměť na fakta).
Hlavní článek: Úloha hipokampu v paměti
Některé důkazy ukazují, že hipokampus se podílí na ukládání a zpracování prostorových informací. Studie na potkanech ukázaly, že neurony v hipokampu mají prostorová vypalovací pole. Tyto buňky se nazývají buňky místa. Některé buňky vzplanou, když se zvíře ocitne na určitém místě, bez ohledu na směr pohybu, zatímco většina je alespoň částečně citlivá na směr hlavy a směr pohybu.
Hlavní článek: Úloha hipokampu v prostorové paměti a navigaci
Hipokampus a deprese
- Úbytek objemu hipokampu,možná odrážející účinky nadměrného množství glukokortikoidů na
neurogenezi, byl zjištěn u skupin lidí s depresí ve srovnání s kontrolními skupinami.
Hlavní článek: Deprese a hipokampus
Historie
Anatom Giulio Cesare Aranzi (asi 1564) poprvé použil termín hipokampus pro označení mozkového orgánu kvůli jeho vizuální podobnosti s mořským koníkem. Tento orgán byl zpočátku spojován spíše s čichem než s jeho známou funkcí při získávání paměti. Na roli hipokampu v paměti upozornil kolem roku 1900 Rus Vladimir Bechtěrev na základě pozorování pacienta s hlubokými poruchami paměti. Po mnoho let však převládal konvenční názor, že hipokampus je stejně jako zbytek limbického systému zodpovědný za emoce.
Na význam hipokampu v paměti upozornil vědce pacient HM. HM trpěl řadou anterográdních a časově odstupňovaných retrográdních poruch paměti (tyto poruchy jsou předmětem filmu Memento) poté, co mu byly oboustranně odstraněny různé struktury mediálního temporálního laloku (včetně oboustranné ablace hipokampů), aby se zmírnily časté epileptické záchvaty. Zvláště důležité je, že HM byl stále schopen učit se procedurální úkoly (které jsou spojeny se striatem) a měl nadprůměrné IQ. HM prokázal nápadnou jednodisociaci mezi inteligencí a deklarativní pamětí. Relativní velikost hipokampální formace ve vztahu k celkovému objemu mozku je často zachována u většiny savčích druhů. Přesto bylo zjištěno, že tyto oblasti jsou u kytovců relativně hypotrofické.
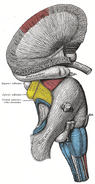
Anatomie
Hlavní článek: Anatomie hipokampu
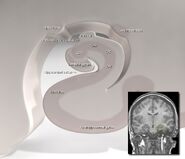
Human hippocampus.
Nissl-stained coronal section of the brain of a macaque monkey, showing hippocampus (circled). Zdroj: brainmaps.org
Anatomicky je hipokampus rozvinutím okraje mozkové kůry. Lze jej rozlišit jako zónu, kde se kůra zužuje do jediné vrstvy velmi hustě uspořádaných neuronů, která se stáčí do těsného esovitého tvaru. Struktury, které lemují okraj kůry, tvoří tzv. limbický systém (latinsky limbus = hranice): patří sem hipokampus, cingulární kůra, čichová kůra a amygdala. Paul MacLean kdysi v rámci své teorie trojjediného mozku navrhl, že limbické struktury tvoří nervový základ emocí. Většina neurovědců však již nevěří, že koncept jednotného “limbického systému” je platný.
Hipokampus jako celek nakonec vypadá jako zakřivená trubice, která byla různě analogizována k mořskému koníkovi, beranímu rohu (Cornu Ammonis) nebo banánu.Toto obecné uspořádání platí pro celou škálu savčích druhů, od ježka po člověka, i když detaily se liší. U potkana vypadají dva hipokampy překvapivě jako dvojice banánů spojených u stopky. U lidských nebo opičích mozků je část hipokampu dole, poblíž báze spánkového laloku, mnohem širší než část nahoře. Jedním z důsledků této složitégeometrie je, že příčné řezy hipokampem mohou vykazovat matoucí rozmanitost tvarů v závislosti na úhlu a umístění řezu.
Nejsilnější spojení hipokampu je s entorhinální kůrou (EC), která leží vedle něj ve spánkovém laloku. Povrchové vrstvy EC poskytují hipokampu nejpočetnější vstupy a hluboké vrstvy EC přijímají nejpočetnější výstupy. EC je zase silně a recipročně propojen s mnoha dalšími částmi mozkové kůry. Hipokampus také dostává velmi důležitou projekci z oblasti mediálního septa. Destrukce septální oblasti ruší hipokampální theta rytmus a vážně narušuje některé typy paměti. (Předpokládá se, že takzvané “date rape” drogy mají své amnestické účinky přinejmenším částečně díky antagonizaci cholinergní projekce z mediálního septa do hipokampu.)
Fyziologie
Příklady EEG potkaního hipokampu a neuronální aktivity CA1 v režimech theta (bdění/chování) a LIA (pomalé vlny spánku). Každý graf zobrazuje 20 sekund dat, přičemž nahoře je stopa hipokampálního EEG, uprostřed rastry hrotů ze 40 současně zaznamenaných pyramidových buněk CA1 (každá linie rastru představuje jinou buňku) a dole graf rychlosti běhu. Horní graf představuje časový úsek, během něhož potkan aktivně hledal rozptýlené pelety s potravou. V případě spodního grafu krysa spala.
Hipokampus vykazuje dva hlavní “režimy” aktivity, z nichž každý je spojens odlišným vzorcem vln EEG a aktivitou nervové populace. tyto režimy jsou pojmenovány podle vzorců EEG, které jsou s nimi spojeny: theta a velká nepravidelná aktivita (LIA). Zde jsou některé z jejich hlavních charakteristik u potkana, zvířete, které bylo nejrozsáhleji studováno:
Mód theta se objevuje během stavů aktivního, bdělého chování (zejména lokomoce) a také během spánku REM (snění). Vtheta režimu dominují v EEG velké pravidelné vlny s frekvenčním rozsahem 6-9 Hz a hlavní skupiny hipokampálních neuronů(pyramidové buňky a granulární buňky) vykazují řídkou populační aktivitu,což znamená, že v jakémkoli krátkém časovém intervalu je velká většina buněk tichá, zatímco malý zbývající zlomek střílí relativně vysokou rychlostí, u nejaktivnějších z nich až 50 hrotů za jednu sekundu. Aktivní buňka obvykle zůstává aktivní půl sekundy až několik sekund. Jak se krysa chová, aktivní buňky utichají a nové buňky se stávají aktivními, ale celkové procento aktivních buněk zůstává víceméně konstantní. V mnoha situacích je aktivita buněk do značné míry určena prostorovou polohou zvířete,ale zjevně ji ovlivňují i další proměnné chování.
Režim LIA se objevuje během pomalých vln (bez snění) spánku a také během stavů bdělé nehybnosti, jako je odpočinek nebo jídlo. V režimu LIA dominují v EEG ostré vlny, což jsou náhodně načasované velké výchylky EEG signálu trvající 200-300 msec. Tyto ostré vlny také určují vzorce nervové aktivity populace. Mezi nimi jsou pyramidové buňky a granulární buňky velmiklidné (ale ne tiché). Během ostré vlny může až 5-10 % populace emitovat akční potenciály během periody 50 msec; mnoho z těchto buněk emituje nikoliv jeden, ale sérii hrotů.
Tyto dva režimy hipokampální aktivity lze pozorovat u primátů stejně jako u potkanů, s důležitou výjimkou, že u primátů bylo obtížné pozorovat robustní theta rytmicitu hipokampu. Existují však kvalitativně podobné ostré vlny a podobné změny aktivity nervové populace závislé na stavu.
Rytmus theta
Vzhledem k hustému uspořádání nervových vrstev generuje hipokampus jedny z největších signálů EEG ze všech mozkových struktur. V některýchsituacích v EEG převládají pravidelné vlny, které často trvají několik sekund. Tento vzorec EEG je známý jako rytmus theta. Byl to jeden z prvních objevených jevů EEG: první popis pochází od Junga a Kornmullera z roku 1938. Teprve v roce 1954, kdy Green a Arduini publikovali dlouhou a důkladnou studii o theta rytmu u králíků, koček a opic, se zájem o tento jev skutečně rozproudil.Možná hlavně proto, že spojili theta rytmus se vzrušením, což bylo v té době aktuální téma, vyvolala jejich práce záplavu navazujících studií, což vedlo k publikování doslova stovek studií o fyziologii a farmakologii theta v průběhu 50. a 60. let. Navzdory tomuto velmi rozsáhlému množství prací zůstávala řada otázek nezodpovězena, zejména otázka funkce. Ani v současnosti není tato nejkritičtější z otázek dosud přesvědčivě zodpovězena.
Rytmicita theta je velmi zřejmá u králíků a hlodavců a je také zřetelně přítomna u koček a psů. Zda lze theta pozorovat i u primátů, je zapeklitá otázka. Green a Arduini zaznamenali u opic pouze velmi krátké výbuchy spíše nepravidelné rytmicity a většina pozdějších studií zaznamenala jen o málo více. Rozdíly v metodice však ztěžují vyvození přesvědčivých závěrů.
U potkanů (zvířata, která byla zdaleka nejrozsáhleji studována) je theta pozorována především za dvou podmínek: za prvé, když zvíře chodí nebo nějakým jiným způsobem aktivně komunikuje se svým okolím; za druhé, během spánku REM. Frekvence se zvyšuje v závislosti na rychlosti běhu, začíná přibližně na 6,5 Hz na nízkém konci a zvyšuje se na přibližně 9 Hz na vysokém konci, ačkoli vyšší frekvence jsou někdy pozorovány u dramatických pohybů, jako jsou skoky přes velké mezery. U jiných, větších druhů zvířat jsou frekvence theta obecně o něco nižší. Zdá se, že závislost chování se také liší podle druhu: u koček a králíků je theta často pozorována během stavů nehybné bdělosti. To bylo zaznamenáno i u potkanů, ale pouze tehdy, když jsou silně vyděšeni.
Theta se neomezuje pouze na hipokampus. U potkanů ji lzepozorovat v mnoha částech mozku, včetně téměř všech, kterés hipokampem silněinteragují. Předpokládá se, že pacemaker tohoto rytmu leží v oblasti mediálního septa: tato oblast se promítá do všech oblastí, které vykazují rytmicitu theta, a její zničení eliminuje theta v celém mozku. (Může existovat jedna výjimka, malá oblast v hypotalamu zvaná supramamilární jádro, která se zdá být v některých situacích schopna udržet theta nezávisle na septu.)
Funkce theta, za předpokladu, že nějakou má, nebyla dosud přesvědčivě vysvětlena, ačkoli bylo navrženo mnoho teorií.Nejpopulárnějším trendem je spojovat ji s učením a pamětí. je dobře známo, že léze středního septa – centrálního uzlu systému theta – způsobují závažné poruchy paměti. střední septum je však více než jen regulátorem theta, je také hlavním zdrojem cholinergních projekcí do hipokampu. Nebylo zjištěno, že by léze septa působily specificky eliminací theta.
Ostré vlny
Během spánku nebo během bdělého stavu, kdy zvíře odpočívá nebo se jinak nezabývá svým okolím, vykazuje hipokampální EEG vzor nepravidelných pomalých vln, které mají o něco větší amplitudu než vlny theta. Tento vzorec je občas přerušován velkými nárazy, které se nazývají ostré vlny. Tyto události jsou spojeny s výbuchy hrotové aktivity trvajícími 50-100 msec v pyramidových buňkách CA3 aCA1. Jsou také spojeny s krátkodobými vysokofrekvenčními EEGoscilacemi zvanými “ripples”. Vlnění s frekvencí v rozmezí150-200 Hz u potkanů lze obvykle detekovat pouze elektrodami umístěnými buď uvnitř buněčné vrstvy CA1, nebo v její těsné blízkosti. Naproti tomu elektrody umístěné kdekoli uvnitř hipokampu nebo dokonce v sousedních mozkových strukturách často zachytí ostré vlny jako velké pomalé výchylky EEG, které trvají 200-400 ms.
U potkanů jsou ostré vlny nejsilnější během spánku, kdy se objevují průměrnou rychlostí kolem 1 za sekundu, ale ve velmi nepravidelném časovém vzorci. Ostré vlny se vyskytují také během neaktivních bdělých stavů, ale tehdy jsou méně časté a obvykle menší. Ostré vlny byly pozorovány také v lidském spánkovém laloku a v opičím hipokampu.U opic jsou ostré vlny poměrně robustní, ale nevyskytují se zdaleka tak často jako u potkanů.
Jedním z nejzajímavějších aspektů ostrých vln je, že zřejmě souvisejí s pamětí. Wilson a McNaughton v roce 1994 a v řadě pozdějších studií uvádějí, že když mají hipokampální place buňky překrývající se prostorová pole střelby (a proto často střílejí téměř simultánně), mají tendenci vykazovat korelovanou aktivitu během spánku následujícího po behaviorálním sezení. Bylo zjištěno, že toto zesílení korelace, obecně známé jako reaktivace, je omezeno hlavně na ostré vlny. Bylo navrženo, že ostré vlny jsou ve skutečnosti reaktivací vzorců nervové aktivity, které byly zapamatovány během chování, což je způsobeno posílením synaptických spojenív rámci hipokampu. Tato myšlenka tvoří klíčovou složku teorie “dvoufázové paměti”, kterou zastává Buzsaki a další a která předpokládá, že vzpomínky jsou uloženy v hipokampu během chování a později jsou přeneseny do neokortexu během spánku:ostré vlny mají pohánět hebbovské synaptické změny v theneokortikálních cílech hipokampálních výstupních drah.
Evoluce
Hipokampus má obecně podobný vzhled napříč celou řadou druhů savců, od bazálních, jako je ježek, až po ty “nejvyspělejší”, jako je člověk. poměr velikosti hipokampu k velikosti těla se široce zvyšuje a u primátů je přibližně dvakrát větší než u ježka. Nezvyšuje se však ani zdaleka tak rychle jako poměr velikosti neokortexu k velikosti těla. hipokampus tedy zabírá mnohem větší objem kůry u hlodavců než u primátů.
Existuje také obecný vztah mezi velikostí hipokampu a prostorovou pamětí: při porovnávání mezi podobnými druhy mají ty, které mají větší schopnost prostorové paměti, obvykle větší objem hipokampu. Tento vztah se vztahuje i na rozdíly mezi pohlavími: u druhů, kde samci a samice vykazují výrazné rozdíly ve schopnosti prostorové paměti, mají tendenci vykazovat i odpovídající rozdíly v objemu hipokampu
Nesavčí druhy nemají strukturu mozku, která by se podobala savčímu hipokampu, ale mají takovou, která je mu považována za homologní. Hipokampus, jak bylo uvedeno výše,je v podstatě mediální okraj mozkové kůry. Pouze savci mají plně vyvinutou mozkovou kůru, ale struktura, ze které se vyvinul, zvaná pallium, je přítomna u všech obratlovců, dokonce i u těch nejprimitivnějších, jako je mihule nebo hagus. Pallium se obvykle dělí na tři zóny: mediální, laterální a dorzální. Mediální pallium tvořípředchůdce hipokampu. Vizuálně se hipokampu nepodobá, protože jeho vrstvy nejsou deformovány do tvaru písmene S ani nejsou obaleny dentátovým gyrem, ale na jeho homologii poukazuje silná chemická a funkční příbuznost. Nyní existují důkazy, že tyto stuktury podobné hipokampu se podílejí na prostorovém poznávání u ptáků, plazů a ryb.
U ptáků je shoda dostatečně prokázána, takže většina anatomů označuje mediální palliovou zónu jako “ptačíhipokampus”. Řada druhů ptáků má silné prostorové schopnosti,zejména ti, kteří skrývají potravu. Existují důkazy, že ptáci, kteří schovávají potravu, mají větší hipokampus než jiné druhy ptáků a že poškození hipokampu způsobuje poruchy prostorové paměti.
U ryb je to složitější. U teleostovitých ryb (které tvoří velkou většinu existujících druhů) je přední mozek ve srovnání s ostatními druhy obratlovců podivnědeformovaný. Většina neuroanatomů se domnívá, že přední mozek teleostů je v podstatě zkroucený, jako ponožka obrácená naruby, takže struktury, které u většiny obratlovců leží uvnitř, vedle komor, se u teleostů nacházejí na vnější straně, a naopak. Jedním z důsledků toho je, že se předpokládá, že mediální pallium (“hipokampální” zóna) typického obratlovce odpovídá laterálnímu palliu typické ryby. Experimentálně bylo prokázáno, že některé druhy ryb (zejména zlaté ryby) mají silnou schopnost prostorové paměti, a dokonce si vytvářejí “kognitivní mapy” oblastí, které obývají. Existují důkazy, že poškození laterálního palia narušuje prostorovou paměť (navigace na dlouhé vzdálenosti, jako je například navádění lososů, však zřejmě závisí na jiných mechanismech).
Zdá se tedy, že úloha hipokampální oblasti v navigaci začíná daleko zpět v evoluci obratlovců a předchází rozdělení, k němuž došlo před stovkami milionů let. Zatím není známo, zda mediální pallium hraje podobnou roli u ještě primitivnějších obratlovců, jako jsou žraloci a rejnoci, nebo dokonce mihule a langusty. Některé druhy hmyzu a měkkýšů, jako jsou chobotnice, mají také silné schopnosti prostorového učení a navigace, ale zdá se, že fungují jinak než prostorový systém savců, takže zatím není dobrý důvod domnívat se, že mají společný evoluční původ; neexistuje ani dostatečná podobnost ve struktuře mozku, aby bylo možné u těchto druhů identifikovat něco podobného “hipokampu”.
Viz také
- Amygdalohipokampektomie
- Mediální svazek předního mozku
- Septal. jádra
Odkazy & Bibliografie
Klíčové texty
Knihy
- Aboitiz, F, Morales D, Montiel J (2003). Evoluční původ izokortexu savců: Na cestě k integrovanému vývojovému a funkčnímu přístupu. Behavior. Brain Sciences 26: 535-552.
- Amaral, D; Lavenex P (2006). “Ch 3. Hippocampal Neuroanatomy” Andersen P, Morris R, Amaral D, Bliss T, O’Keefe J The Hippocampus Book, Oxford University Press.
- Broglio, C, Gómez A, Durán E, Ocaña FM, Jiménez-Moya F, Rodríguez F, Salas C (2002). Znaky společného plánu předního mozku obratlovců: Specializované palliální oblasti pro prostorovou, časovou a emoční paměť u aktinopterygií. Brain Res. Bull. 57: 397-399.
- Buzsáki, G (2002). Theta oscilace v hipokampu. Neuron 33: 325-340.
- Buzsáki, G (2006). Rhythms of the Brain.
- Cantero, JL, Atienza M, Stickgold R, Kahana MJ, Madsen JR, Kocsis B (2003). Oscilace theta v závislosti na spánku v lidském hipokampu a neokortexu. J. Neurosci. 23: 10897-10903.
- Colombo, M, Broadbent N (2000). Je ptačí hipokampus funkčním homologem hipokampu savců? Neurosci. Biobehav. Rev. 24: 465-484.
- Eichnbaum, H; Cohen NJ (1993). Memory, Amnesia, and the Hippocampal System, MIT Press.
- Ekstrom, AD, Kahana MJ, Caplan JB, Fields TA, Isham EA, Newman EL, Fried I (2003). Buněčné sítě, které jsou základem lidské prostorové navigace. Nature 425: 184-188.
- Gray, JA; McNaughton N (2000). The Neuropsychology of Anxiety [Neuropsychologie úzkosti]: An Enquiry into the Functions of the Septo-Hippocampal System, Oxford University Press.
- Green, JD, Arduini AA (1954). Hipokampální elektrická aktivita při vzrušení. J. Neurophysiol. 17: 533-557.
- Jacobs, LF, Gaulin SJ, Sherry DF, Hoffman GE (1990). Evoluce prostorového poznávání: pohlavně specifické vzorce prostorového chování předpovídají velikost hipokampu. PNAS 87: 6349-6352.
- Jacobs, LF (2003). Evoluce kognitivní mapy. Brain Behav. Evol. 62: 128-139.
- Kirk, IJ, McNaughton N (1991). Supramammillary cell firing and hippocampal rhythmical slow activity [Vypalování supramammillárních buněk a rytmická pomalá aktivita hipokampu]. Neuroreport 11: 723-725.
- Maguire, EA, Gadian DG, Johnsrude IS, Good CD, Ashburner J, Frackowiak RS, Frith CD (2000). Strukturální změny související s navigací v hipokampu taxikářů. PNAS 97: 4398-4403.
- McNaughton, BL, Battaglia FP, Jensen O, Moser EI, Moser MB (2006). Path integration and the neural basis of the ‘cognitive map’ (Integrace cest a nervový základ “kognitivní mapy”). Nat. Rev. Neurosci. 7: 663-678.
- Moser, EI, Kropf E, Moser M-B (2008). Place Cells, Grid Cells, and the Brain’s Spatial Representation System (Místní buňky, mřížkové buňky a systém prostorové reprezentace mozku). Ann. Rev. Neurosci. 31.
- Nieuwenhuys, R (1982). An Overview of the Organization of the Brain of Actinopterygian Fishes [Přehled organizace mozku aktinopterygních ryb]. Am. Zool. 22: 287-310.
- O’Kane, G, Kensinger EA, Corkin S (2004). Důkaz sémantického učení u hluboké amnézie: An investigation with patient H.M. (Vyšetřování s pacientem H.M.). Hippocampus 14: 417-425.
- O’Keefe, J; Nadel L (1978). The Hippocampus as a Cognitive Map, Oxford University Press.
- Portavella, M, Vargas JP, Torres B, Salas C (2002). Účinky lézí telencefalického patra na prostorové, časové a emoční učení u zlatých rybek. Brain Res. Bull. 57: 397-399.
- Rodríguez, F, Lópeza JC, Vargasa JP, Broglioa C, Gómeza Y, Salas C (2002). Prostorová paměť a hipokampální pallium v průběhu evoluce obratlovců: poznatky z plazů a teleostovitých ryb. Brain Res. Bull. 57: 499-503.
- Sainsbury, RS, Heynen A, Montoya CP (1987). Behaviorální koreláty hipokampální theta typu 2 u potkana. Physiol. Behavior. 39: 513-519.
- Scoville, WB, Milner B (1957). Loss of Recent Memory After Bilateral Hippocampal Lesions [Ztráta recentní paměti po oboustranné lézi hipokampu]. J. Neurol. Neurosurg. Psych. 20: 11-21.
- Shettleworth, SJ (2003). Memory and Hippocampal Specialization in Food-Storing Birds [Paměť a hipokampální specializace u ptáků ukládajících potravu]: Výzvy pro výzkum srovnávacího poznávání. Brain Behav. Evol. 62: 108-116.
- Skaggs, WE, McNaughton BL, Wilson MA, Barnes CA (1996). Theta fáze precese v hipokampálních neuronálních populacích a komprese časových sekvencí. Hippocampus 6: 149-176.
- Squire, LR (1992). Paměť a hipokampus: syntéza poznatků získaných u potkanů, opic a lidí. Psych. Rev. 99: 195-231.
- Squire, LR; Schacter DL (2002). The Neuropsychology of Memory, Guilford Press.
- Vanderwolf, CH (1969). Hipokampální elektrická aktivita a dobrovolný pohyb u potkana. EEG & Clin. Neurophysiol. 26: 407-418.
- Vargas, JP, Bingman VP, Portavella M, López JC (2006). Telencephalon a geometrický prostor u zlatých rybek. Eur. J. Neurosci. 24: 2870-2878.
- West, MJ (1990). Stereologické studie hipokampu: srovnání hipokampálního členění různých druhů včetně ježků, laboratorních hlodavců, divokých myší a lidí. Prog. Brain Res. 83: 13-36.
Dokumenty
- Amaral DG a Cowan WM. 1980. Subkortikální aferenty k hipokampální formaci u opice. Journal of Comparative Neurology. Feb 15; 189(4):573-91.
- Duvernoy, H. (2005) The Human Hippocampus, 3. vydání, Berlin: Springer-Verlag.
Další materiály
Dokumenty
- Google Scholar
- http://www.psycheducation.org/emotion/hippocampus.htm Velké prohlídky mozku!!!
- BrainMaps na UCDavis hippocampus
- Artificial Hippocampus
- BrainInfo na University of Washington hier-164
- John O’Keefe & Lynn Nadel (1978) The Hippocampus as a Cognitive Map , Oxford University Press. Plný text
Další obrázky
Přidat fotografii do této galerie
Lidský mozek: Limbický systém
Amygdala – Cingulate gyrus – Fornicate gyrus – Hippocampus – Hypothalamus – Mammillary body – Nucleus accumbens – Orbitofrontální kůra – Parahippocampální gyrus
|
v-d-e
Lidský mozek: přední mozek (mozková kůra – mozková kůra – mozkové hemisféry, šedá hmota) (TA A14.1.09.002-240, 301-320, GA 9.818-826) |
|||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Frontální lalok |
|
||||||||||||||||
| Parietální lalok |
|
Superolaterální
|
Mediální/dolní
|
obě
|
|||||||||||||
| Occipitální lalok |
|
Superolaterální
|
Medial/inferior
|
||||||||||||||
| Temporální lalok |
|
Superolaterální
|
Mediální/dolní
|
||||||||||||||
| Interlobární sulci/fissures |
. |
Superolaterální
|
Mediální/inferiorní
|
||||||||||||||
| Limbický lalok |
|
Ostatní
|
|||||||||||||||
| Insulární lalok |
Dlouhý gyrus insulární – Krátké gyry insulární – Kruhový sulcus insulární |
||||||||||||||||
| obecný |
Operculum – Póly mozkových hemisfér |
||||||||||||||||
Některé kategorizace jsou přibližné, a některé Brodmannovy oblasti zahrnují gyry.
|
|||||||||||||||||
Tato stránka využívá obsah s licencí Creative Commons z Wikipedie (zobrazit autory).