- Introduction
- Módszerek
- Elterjedési adatok és biomosztályozás
- Fajösszetétel a biomok között
- Trait-adatok
- Trait Hypervolumes Measurement
- Funkcionális megkülönböztethetőség és elterjedtség
- Funkcionális tér és biomok hasonlósága
- Eredmények
- Diszkusszió
- Tudásbeli hiányosságok
- A biomokon belüli közös minták
- Bioomok közötti összehasonlítások
- Következtetések
- Adatok elérhetősége
- A szerzők hozzájárulása
- Finanszírozás
- Kérdések összeférhetetlenségi nyilatkozata
- Köszönet
- Kiegészítő anyagok
Introduction
Ecologists and biogeographers organizizing land plant biodiversity into climatically-determinated biomes, with physiognomies characterized by the growth forms and functional traits of the dominant species (Moncrieff et al., 2016). A biomok koncepciójának valóban hosszú története van, amely Carl Ludwig Willdenowig és Alexander von Humboldtig nyúlik vissza. Willdenow felismerte, hogy a hasonló éghajlatok hasonló növényzeti formákat támogatnak, Humboldt pedig megfigyelte a növények elterjedése, a fiziognómia és a környezeti tényezők közötti széles körű összefüggést. Összességében a biom koncepció azt a feltételezést tükrözi, hogy a hasonló környezeti nyomások hasonló funkcionális tulajdonságokkal rendelkező fajokat választanak ki, függetlenül azok evolúciós történetétől. Ugyanakkor a Föld ma élő biomjai bizonyos mértékig filogenetikai szempontból elkülönülnek egymástól, és a jellemző fajok nagy része/többsége olyan specifikus vonalból származik, amelyek nemcsak az éghajlati szelekcióhoz, hanem olyan további nyomásokhoz, mint a tűz vagy a megafélék (Woodward et al., 2004; Pennington et al., 2006; Donoghue és Edwards, 2014) való kulcsfontosságú alkalmazkodást mutatnak. Mivel a biomok a Föld növényzetének széles körű szabályszerűségeit képviselik, a biomok közötti funkcionális különbségek megértése kritikusan fontos a globális szénciklus és a földi rendszer működésének modellezéséhez, beleértve az antropogén globális változásokra adott válaszokat is (Bonan et al., 2012; van Bodegom et al., 2014; Xia et al., 2015).
Az elmúlt két évtizedben gyorsan nőtt a növényi biodiverzitás kontinentális léptékű funkcionális dimenziójának megértése (Swenson et al., 2012; Lamanna et al., 2014; Šímová et al., 2018). A kiterjedt adatszintézisre és -elemzésre irányuló erőfeszítések egy korlátozott funkcionális tulajdonságteret határoztak meg, amely magában foglalja a teljes növényi méret, a levélgazdaságosság és a hidraulika variációját, az úgynevezett funkcionális tulajdonságspektrumokat (Wright et al., 2004; Díaz et al., 2016). A funkcionális tulajdonságspektrumok olyan fiziológiai és ökológiai kompromisszumokat határoznak meg, amelyek meghatározzák a növények életmód-stratégiáit (Westoby, 1998; Reich et al., 2003; Craine, 2009), és amelyek hatással vannak a közösségek összeállására (Kraft et al., 2008), az ökoszisztémák működésére (Lavorel és Garnier, 2002; Garnier et al., 2004; Kerkhoff et al., 2005; Cornwell et al., 2008), sőt az evolúció sebességére is (Smith és Donoghue, 2008; Smith és Beaulieu, 2009). A növényi funkcionális tulajdonságspektrumok általánossága és hasznossága felgyorsította beépítésüket a növényzet globális eloszlásának (van Bodegom et al., 2014), biogeokémiai körforgásának (Bonan et al., 2012) és ökoszisztéma-szolgáltatásainak (Díaz et al., 2007; Cadotte et al., 2011; Lavorel et al., 2011) modelljeibe. Kevés tanulmány vizsgálta azonban a funkcionális diverzitás átfogó mérését biom léptékben mind a domináns, mind az alárendelt fajok és életformák tekintetében.
Két ellentétes előrejelzés létezik arra vonatkozóan, hogy a funkcionális diverzitás hogyan változik az éghajlati és fiziognómiai szempontból különböző biomok között. Egyrészt, ha a biomok biológiai sokfélesége a rendelkezésre álló ökológiai niche-tér változatosságát tükrözi, akkor a taxonómiailag kevésbé változatos biomoknak a változatosabb biomok által elfoglalt funkcionális tér kisebb, nagyrészt egymásba ágyazott részhalmazát kellene képviselniük. Másrészt, a növényi szerkezet és funkció alapvető kompromisszumainak globális jellege és a fajok között hasonlóan ható hasonló szelekciós nyomások miatt a különböző környezeteket elfoglaló együttesek valójában a tulajdonságtér hasonló területein osztozhatnak (Reich et al., 2003; Wright et al., 2004; Díaz et al., 2016). A legújabb tanulmányok megerősítik, hogy az éghajlat és a funkcionális és taxonómiai diverzitás (azaz a fajgazdagság) közötti kapcsolatok összetettek és skálafüggőek. Egyes esetekben a funkcionális diverzitás szorosan követi az éghajlati gradienseket, alacsonyabb funkcionális diverzitással a változékonyabb és szélsőségesebb körülmények között (Swenson et al., 2012; de la Riva et al., 2018), és a taxonómiai gazdagság növekedésével bővül a funkcionális tér mennyisége a helyi együttesekben (Lamanna et al., 2014; Li et al., 2018). A regionális léptékű fajkészletek elemzései azonban arra utalnak, hogy a fajgazdagság nagymértékű változásai minimális hatással járhatnak a funkcionális diverzitásra (Lamanna et al., 2014; Šímová et al., 2015). Ráadásul a taxonómiai és funkcionális diverzitás éghajlatra adott válaszai különbözhetnek a főbb növényi növekedési formák között (Šímová et al., 2018), és a különböző növekedési formák relatív diverzitása jelentősen változhat a különböző biomokat meghatározó éghajlati gradiensek mentén (Engemann et al., 2016).
A funkcionális diverzitás kontinentális és globális léptékű elemzése terén elért eredményeket részben a biológiai sokféleség taxonómiai, elterjedési, filogenetikai és funkcionális szempontjainak katalogizálásában mutatkozó adathiányok korlátozzák (Hortal et al., 2015). Továbbá, még a korlátozottan rendelkezésre álló adatok mellett is jelentős informatikai kihívásokat jelent a fajok földrajzi elterjedését, funkcionális jellemzőit és filogenetikai kapcsolatait leíró nagy, eltérő adathalmazok szabványosítása és integrálása (Violle et al., 2014). Itt a növényfajok funkcionális diverzitásának eloszlását vizsgáljuk Észak- és Dél-Amerika biomjai között a Botanical Information and Ecology Network adatbázis (BIEN; Enquist et al., 2016; Maitner et al., 2018) segítségével, amely >100 000 szárazföldi növényfaj eloszlási és funkcionális tulajdonságokra vonatkozó információit állítja össze. Konkrétabban a funkcionális diverzitás és a biomok megkülönböztethetőségének változatosságát vizsgáljuk a szárazföldi növényfajok 100 × 100 km-es rácshálós cellák méretarányú elterjedési térképei és az ökológiai stratégiák változatosságának fő tengelyeit tükröző hat funkcionális tulajdonság átfogó adatkészlete alapján.
A tanulmányunk céljai hármasak. Először is, hogy rávilágítsunk a tartós adathiányra, dokumentáljuk a biomok funkcionális sokféleségét és megkülönböztethetőségét jellemző rendelkezésre álló adatok mértékét. Másodszor, a rendelkezésre álló adatok alapján jellemezzük a funkcionális sokféleség eloszlását a biomokon belül mind a domináns, mind az alárendelt növekedési formák között. Ezek az elemzések lehetővé teszik számunkra, hogy jobban számszerűsítsük egy biom funkcionális megkülönböztethetőségét azáltal, hogy azonosítjuk a biomon belül a legelterjedtebb fajok leggyakoribb funkcionális stratégiáit. Harmadszor, a funkcionális tulajdonságok terében a többdimenziós hipervolumenek átfedése alapján megkérdezzük, hogy a biomokat valóban funkcionálisan elkülönülő fajgyűjtemények jellemzik-e.
Módszerek
Elterjedési adatok és biomosztályozás
A több forrásból összeállított előfordulási adathalmazokra jellemző mintavételi torzítás hatásainak csökkentése érdekében az Észak- és Dél-Amerikában elterjedt 88 417 növényfajra vonatkozó BIEN 2.0 elterjedési térképeket használtuk (Goldsmith et al, 2016). A BIEN adatbázis a herbáriumi példányokból és a vegetációs parcellák leltározásából származó szabványosított növénymegfigyeléseket integrálja. A BIEN-adatbázisokban szereplő fajok elterjedési terület-térképeit háromféle megközelítés egyikével becsülték meg, attól függően, hogy az egyes fajokról milyen előfordulások állnak rendelkezésre. A csak egy vagy két előfordulással rendelkező fajok esetében (a fajok kb. 35%-a) a földrajzi elterjedési területet az egyes adatpontokat körülvevő 75 000 km2 -es négyzet alakú területként határoztuk meg. A három vagy négy előfordulással rendelkező fajok (a fajok kb. 15%-a) földrajzi hatókörét egy konvex héj segítségével határoztuk meg. Végül a legalább öt előfordulással rendelkező fajok elterjedési térképeit a Maxent fajeloszlás-modellező algoritmus segítségével kaptuk meg, 19 éghajlati réteget használva előrejelző változóként és 19 térbeli sajátvektort szűrőként a modellek előrejelzéseinek korlátozása érdekében (lásd Goldsmith et al, 2016 az elterjedési térképek módszertanának további részleteiért).
A BIEN 2.0 növényfaj-térképeket egy 100 × 100 km-es rácshálós térképre helyeztük rá Lambert Azimuthal Equal Area vetülettel, hogy minden rácscellára megkapjuk a fajok jelenlét/hiány mátrixát. Az Olson et al. (2001) biom osztályozása alapján minden mátrixrácscellát a biomok egyik kategóriájába soroltunk. A későbbi elemzésekhez szükséges számítási korlátok miatt egyes biomokat az éghajlati és vegetációs hasonlóságok, valamint a szakirodalom alapján egyesítettünk, hogy az 1. táblázatban leírt széles körű besorolást kapjuk (kivéve a belvizek, a szikla- és jégrétegek, valamint a mangrovék). A Chaco és a Caatinga ökorégiókat a Xeric Woodlands és a Dry forest (száraz erdők) kategóriába soroltuk, Prado és Gibbs (1993), Pennington et al. (2000), Banda et al. (2016), Silva de Miranda et al. (2018) alapján.
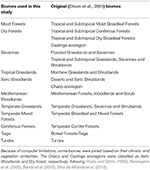
Táblázat 1. táblázat. Az ebben a tanulmányban elfogadott biomok osztályozásának és az ezzel egyenértékű (Olson et al., 2001) biomok osztályozásának áttekintése (kivéve a belvizek, a szikla és jég, valamint a mangrove).
Fajösszetétel a biomok között
A funkcionális tulajdonságtérben a biomok közötti eltérések megértéséhez először a növényi fajösszetétel közötti különbségeket vizsgáltuk. Az egyes rácscellák és biomok fajlistája alapján az egyes biomok jellemző fajait úgy határoztuk meg, hogy azok a fajok, amelyek elterjedési területük legnagyobb hányadát az adott biomban találhatók. Az adott biomban földrajzilag uralkodó fajok listáját összehasonlítottuk a többi biomban előforduló fajok listájával. Ez a páros összehasonlítás egyszerű módon lehetővé teszi a biomok közötti irányított taxonómiai átfedés megállapítását (azaz egy biom domináns fajainak egy másik biommal közös arányát). A 88 417, elérhető elterjedési területi térképekkel rendelkező faj közül 44 899 faj elterjedési területe egynél több biomra terjed ki, és 43 518 faj endemikus egy adott biomban.
Trait-adatok
Az Újvilágban élő növényfajok összes, a BIEN 3.0 adatállományban (lekérdezés 2018. február 7-én) elérhető tulajdonságinformációját kinyertük, így összesen 80 405 fajszintű tulajdonságmegfigyelést kaptunk. Ezután megszűrtük az információkat hat funkcionális tulajdonságra: maximális növénymagasság (m), magtömeg (mg), fasűrűség (mg/cm3), fajlagos levélfelület SLA (cm2/g) és levélfoszfor- és levélnitrogén-koncentráció egységnyi tömegre vetítve (mg/g). A szűrés után összesen 18 192 fajszintű megfigyelés maradt. Ezek közül azt a 8 820 fajból álló részhalmazt használtuk fel a további elemzésekhez, amely mind a tartománytérképekkel, mind a tulajdonságinformációkkal rendelkezett.
A funkcionális tulajdonságtér becsléséhez az egyes biomok esetében teljes tulajdonságadatokra volt szükségünk. Ezért a hiányzó tulajdonságadatokat filogenetikai úton imputáltuk (Bruggeman et al., 2009; Penone et al., 2014; Swenson, 2014; Swenson et al., 2017) az “Rphylopars” v 0.2.9 R-csomag (Goolsby et al., 2017) és a Smith és Brown (2018) által nemrég közzétett, a magról kelő növények filogenetikáját használva alapként. A filogenetikai imputáció a funkcionális tulajdonságok adathalmazaiban hiányzó adatok előrejelzésére szolgáló eszköz, amely azon a feltételezésen alapul, hogy a közeli rokonságban álló fajok általában hasonló tulajdonságértékekkel rendelkeznek (Swenson, 2014). Az ALLBM fát használtuk (azaz a Magallón et al. (2015) által biztosított génbank és az Open Tree of life taxonok gerinccel), mivel ez maximalizálta a rendelkezésre álló tulajdonság- és elterjedési információkkal rendelkező fajok közötti átfedést. A tulajdonságimputáció után összesen 7842, teljes tulajdonságinformációval és elterjedési térképekkel rendelkező faj maradt a további elemzéshez.
Trait Hypervolumes Measurement
A biomok relatív funkcionális diverzitását a biomsejteken belüli fajkészletekből származó tulajdonsághipervolumes kiszámításával mértük. A számítási korlátok miatt a trait hipervolumeneket az egyes biomok celláinak 20%-ából vett véletlenszerű minta alapján állítottuk össze. Az egyes rácscellákra vonatkozó hipervolumeneket a kivont hat funkcionális tulajdonság és a “hypervolume” R-csomag (Blonder et al., 2014, 2018) segítségével becsültük meg, a Gauss-féle KDE-módszerrel, az alapértelmezett Silverman-sávszélességbecslővel. A magtömeget, a magasságot és a fasűrűséget log-transzformáltuk, míg az SLA-t négyzetgyök-transzformáltuk. Az elemzés előtt minden tulajdonságot skáláztunk és központosítottunk. A hipervolumeneket a standard eltérések (sd) mértékegységében adjuk meg a felhasznált tulajdonságok számának hatványára (azaz sd6).
Funkcionális megkülönböztethetőség és elterjedtség
Mivel a biomokat a domináns növényzetük jellemzi, megvizsgáltuk a fajok földrajzi elterjedtségét és funkcionális megkülönböztethetőségét is a biomokon belül. A fajok elterjedési területének térképeit és a funkcionális tulajdonságokra vonatkozó információkat felhasználva megbecsültük az egyes fajok funkcionális megkülönböztethetőségét és elterjedtségét az egyes biomokban, a Violle et al. (2017) által a funkcionális ritkaság fogalmi keretét követve. A (mért és imputált) tulajdonságok teljes készletét felhasználva először megmértük az euklideszi távolságot a standardizált tulajdonságtérben az összes fajpár között. Ezután kiszámítottuk az egyes fajok funkcionális megkülönböztethetőségét (Di) az egyes bioszférákban, mint egy faj átlagos funkcionális távolságát az adott bioszféra fajkészletén belüli N másik fajhoz képest. A Di-t 0 és 1 között skáláztuk az egyes biomokon belül, az alacsonyabb értékek a funkcionálisan közös (redundáns) fajokat, a magasabb értékek pedig a funkcionálisan megkülönböztető fajokat jelentik az adott biom többi fajához képest. A fajok földrajzi elterjedtségét (Wi) is megbecsültük egy bioszférában, amelyet a faj által egy bioszférán belül elfoglalt rácskockák számának az adott bioszféra összes cellájához viszonyított arányaként mértünk. Az 1 érték azt jelzi, hogy a fókuszfaj a biom által lefedett összes rácshálós cellában jelen van. Mind a funkcionális megkülönböztethetőséget, mind a földrajzi elterjedtséget a “funrar” R-csomag (Grenié et al., 2017) segítségével számoltuk ki.
Mivel nem rendelkezünk a dominancia helyi abundancián vagy biomasszán alapuló átfogó mérőszámokkal, a Di és Wi mérőszámokat használtuk arra, hogy minden egyes biom esetében azonosítsuk a “leggyakoribb” fajokat, mint amelyek mind földrajzi szempontból elterjedtek (Wi > 0,5), mind funkcionálisan hasonlóak (Di < 0,25). Ez utóbbi küszöbértéket használtuk, mivel a funkcionális megkülönböztethetőségi értékek harmadik kvantilisének értéke a biomok között 0,2 és 0,3 között mozgott. A funkcionálisan eltérő fajok és a funkcionálisan hasonló fajok megkülönböztetéséhez a későbbi elemzésekben ezért 0,25-ös értéket használtunk határértékként (azaz, a < 0,25-ös megkülönböztethetőségi értékkel rendelkező fajokat funkcionálisan közösnek vagy redundánsnak tekintettük egy adott biomon belül).
Funkcionális tér és biomok hasonlósága
A biomok hipervolumenei közötti átfedés becsléséhez a Sørensen-féle hasonlósági indexet alkalmaztuk (i) a fajok teljes száma és (ii) az egyes biomok esetében funkcionálisan közösnek és földrajzilag elterjedtnek tekintett fajok listája alapján. A funkcionális tulajdonságok térbeli hasonlóságát a biomok közötti hipervolumenek páronkénti töredékes átfedéseként számoltuk ki. A tört átfedést úgy számították ki, hogy két hipervolumen metszéspontjának térfogatát elosztották az egyesülésük térfogatának kétszeresével. Minden hipervolument a “hypervolume” R-csomag (Blonder et al., 2014, 2018), a Gauss KDE módszerrel, az alapértelmezett Silverman sávszélesség-becslővel.
Eredmények
A modellezett fajterjedelmek metszéspontja alapján jelentős átfedést találtunk a növényfajok összetételében az egyes biomok között (lásd az 1. ábrát és az 1. kiegészítő táblázatot). Ez a taxonómiai átfedés nagyobb a trópusi és a mérsékelt égövi biomokon belül, viszonylag kevés fajon osztozik ez a két éghajlati övezet. Érdekes módon a xerikus erdőterületek mind a trópusi, mind a mérsékelt égövi biomokkal közös fajokkal rendelkeznek. A trópusi nedves erdőkre jellemző fajok magas aránya (83%) ellenére ez a biom nagyszámú fajon osztozik más trópusi biomokkal, mint például a száraz erdőkkel, szavannákkal, száraz erdőkkel és trópusi gyepekkel. A mérsékelt égövi biomok, például a mérsékelt égövi gyepek, a mediterrán erdők, a tajga és a tundra nagyfokú taxonómiai átfedése arra utal, hogy a funkcionális megkülönböztethetőség alacsony lehet, és egyes mérsékelt égövi biomok a fajgazdagabb, funkcionálisan sokszínűbb trópusi biomok szegényebb részhalmazát képviselik.
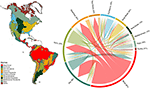
1. ábra. A növényfajok átfedése az Újvilág biomjai között. A százalékos értékek az egy biomban előforduló azon fajok hányadát fejezik ki, amelyek földrajzi elterjedési területük legnagyobb hányadát az adott biomban találjuk, N = 84 413. Az egyes elágazó nyilak alja úgy van elhelyezve, hogy azt a biomot mutassa, amely a faj elterjedési területének legnagyobb részét foglalja magában, míg a szélesség a nyíl hegyénél lévő biomokkal közös fajok számát jelzi. Az alapul szolgáló mátrixot lásd az 1. kiegészítő táblázatban.
Mivel a feltérképezett fajok kevesebb mint 10%-a képviseltette magát, a tulajdonságadatok meglehetősen gyéren álltak rendelkezésre a biomi léptékű fajösszetételeinkre vonatkozóan (2A. ábra). Ráadásul a rendelkezésre álló tulajdonságadatok a tulajdonságok, a növényi kládok és a biomok között jelentős eltéréseket mutattak. Filogenetikai szempontból a levél P és a faanyag sűrűsége különösen gyengén volt reprezentálva, egész kládok esetében nem állt rendelkezésre semmilyen adat. Ennek eredményeképpen a 7 842 fajra vonatkozó tulajdonságértékek körülbelül 2/3-ának imputálására volt szükség, és sok tulajdonságot közeli rokon taxonok hiányában kellett hozzárendelni. Földrajzi szempontból az SLA, a magasság és a magtömeg alulmintázott a trópusokon és a mérsékelt égövi Dél-Amerikában, míg a levél N és a faanyag sűrűsége egyenletesebb a régiók között. A levél P mintázata minden biomban gyenge. Az egyes biomok esetében a sejtek 20%-ának véletlenszerű kiválasztásából álló fajkészletekből létrehozott hipervolumenek főként a trópusi és a mérsékelt égövi/hideg biomok közötti különbségeket mutatnak (2B. ábra). A tulajdonságok hipervolumenei általában nagyobbak és változékonyabbak a trópusi biomokban, a legnagyobb eltérésekkel a nedves és száraz trópusi erdőkben. A mérsékelt égövi/hideg biomok közül a tűlevelű és a mérsékelt égövi vegyes erdők támogatják a legnagyobb hipervolumenű tulajdonságokat. Érdekes módon a xerikus bozótosok a hipervolumenek eloszlása jobban hasonlít a trópusi, mint a mérsékelt égövi biomokéhoz.
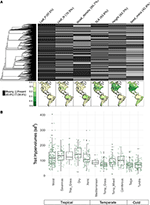
2. ábra. (A) A BIEN 3.0 adatbázisban ismert (szürke) vagy hiányzó (fekete) tulajdonságértékekkel rendelkező fajok aránya. A bal oldali filogenezis megfelel a Smith és Brown (2018) magoncnövények ALLBM-fájának (azaz a GenBank és az Open Tree of life taxonok a Magallón et al. (2015) által megadott gerinccel). A térképek a tulajdonságinformációval rendelkező fajok arányát mutatják a BIEN 2.0 adatbázisban szereplő fajok teljes számához viszonyítva. (B) Az egyes biomokban véletlenszerűen kiválasztott 100 × 100 km-es cellák 20%-ának vonáshypervolumenének eloszlása. A hipervolumeneket a felhasznált tulajdonságok számának hatványa szerinti standard eltérések mértékegységében adjuk meg.
A funkcionális megkülönböztethetőség és a földrajzi eloszlás közötti kapcsolat figyelemre méltóan hasonló az egyes biomok között (3. ábra). Minden biomban a fajok túlnyomó többsége egyszerre földrajzilag korlátozott és funkcionálisan hasonló. Ugyanakkor a funkcionálisan legjellegzetesebb fajok minden biomon belül általában földrajzilag is korlátozottak voltak, és a földrajzilag legelterjedtebb fajok szinte mindig funkcionálisan hasonlóak voltak.
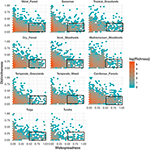
3. ábra. A funkcionális megkülönböztethetőség mintázatai az egyes biomok között. A megkülönböztethetőség azt mutatja, hogy a fajok funkcionálisan mennyire távol vannak egymástól egy biomon belül (azaz a fókuszfaj és az összes többi faj átlagos páronkénti fenotípusos távolsága). Minél nagyobb az érték, annál távolabb van egy faj a biom funkcionális terének középpontjától. A elterjedtség azt méri, hogy egy faj földrajzilag mennyire gyakori egy biomon belül. A 0 érték azt jelzi, hogy egy faj egyetlen biomi cellában van jelen. A bin-értékek a fajok számát jelentik egy logaritmikus skálán. A négyszögek azokat a fajokat ábrázolják, amelyeket az egyes biomokban funkcionálisan hasonlónak és elterjedtnek tekintünk, a megkülönböztethetőség és az elterjedtség 0,25, illetve 0,5 határértékei alapján.
A növekedési formák biomon belüli és biomok közötti arányos eloszlásában jelentős eltéréseket találtunk. A fajszámot tekintve a trópusi biomokban a fás fajok képviselik a domináns növekedési formákat, míg a mérsékelt égövi környezetben a lágyszárú fajok dominálnak (4. ábra). Amikor csak azokat a fajokat vettük figyelembe, amelyek széles körben elterjedtek és funkcionálisan gyakoriak az egyes biomokban, a növekedési formák eloszlása a biomok között megváltozott, különösen a fák, a lágyszárúak és a fűfélék arányában. A trópusi biomokban például a nedves erdők kivételével minden biomban csökkent a fák aránya. A mérsékelt égövi biomokban a fűfélék aránya nőtt, különösen a sarkvidékek felé. Érdekes módon a xerikus erdőkben a növekedési formák eloszlása jobban hasonlít a mérsékelt égövi biomok eloszlásához, ha csak a széles körben elterjedt és funkcionálisan gyakori fajokat vesszük figyelembe.
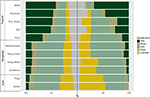
4. ábra. A növekedési formák eloszlása az egyes biomokban (balra) a fajok teljes száma alapján, illetve (jobbra) csak a funkcionálisan hasonló és az adott biomban elterjedt fajok alapján (lásd a 3. ábrát).
A fajok összetételének páronkénti összehasonlítása a biomok között három fő klasztert mutat, amelyek a trópusi, mérsékelt és sarkvidéki éghajlati övezetet képviselik (5A. ábra), ami tükrözi a nagyfokú taxonómiai átfedést e régiókon belül, de nem ezek között (1. ábra). A tulajdonságok hipervolumenének páronkénti összehasonlítása a biomok között az éghajlati övezetek kevésbé egyértelmű klaszterezését mutatja (5B. ábra). Ebben az esetben a xerikus erdők funkcionális tere jelentősen átfedésben van a mérsékelt égövi biomokkal. Ez az elemzés a taigák funkcionális terének a mérsékelt égövi gyepekkel és vegyes erdőkkel való átfedését is feltárja. A tulajdonságok hipertérfogatának a biomok közötti páronkénti összehasonlítása csak a funkcionálisan gyakori és elterjedtnek tekintett fajok felhasználásával a tulajdonságtérben kisebb átfedést mutat az éghajlati övezetek között és azokon belül (5C. ábra). Annak ellenére azonban, hogy a xerikus erdők most a többi trópusi biomával együtt csoportosulnak, ezek az élőhelyek a trópusi gyepekkel együtt nagy átfedést mutatnak a funkcionális térben a mérsékelt égövi biomákkal, például a mediterrán erdőkkel és a mérsékelt égövi gyepekkel.
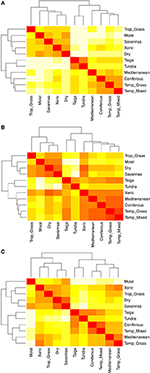
5. ábra. (A) A fajösszetétel páronkénti különbözősége a biomok között. (B) Páros eltérés a tulajdonságok hipervolumenében (1-Sørensen-hasonlóság) a biomok között az összes fajszám alapján. (C) A tulajdonságok hipervolumenének páronkénti különbözősége (1-Sørensen-hasonlóság) a biomok között, csak a funkcionálisan hasonlónak és elterjedtnek tekintett fajok felhasználásával. Minél világosabb a cella, annál nagyobb az eltérés.
Diszkusszió
Elemzéseink három fontos felismerést hoztak a szárazföldi biomok funkcionális szemszögből történő megértéséhez. Először is, megmutatjuk, hogy az elsődleges biodiverzitási adatok összeállításában és szintézisében elért haladás ellenére továbbra is jelentős tudáshiány van, ami korlátozhatja a biomok funkcionális biodiverzitásának számszerűsítésére való képességünket kontinentális és globális léptékben. Másodszor, a rendelkezésre álló adatok elemzése mindazonáltal azt mutatja, hogy Észak- és Dél-Amerika valamennyi biomjának van egy figyelemre méltó közös mintája, amely szerint a földrajzilag legelterjedtebb fajok bármely biomban általában közös funkcionális tulajdonságokkal rendelkeznek, míg a funkcionálisan legjellegzetesebb fajok elterjedése kivétel nélkül korlátozott. Harmadszor, ha csak a széles körben elterjedt és funkcionálisan közös fajokat vesszük figyelembe, a biomok funkcionálisan könnyebben megkülönböztethetők, és úgy tűnik, hogy a biomok közötti különbségek mintázatai az éghajlat és a növényi funkcionális réstér közötti megfelelést tükrözik. Eredményeink összességében azt sugallják, hogy bár a biomok funkcionális sokféleségének vizsgálata még kialakulóban van, a terület további fejlődése valószínűleg az evolúciót, a biogeográfiát, a közösségek összeállását és az ökoszisztémafunkciókat összekötő betekintést fog nyújtani.
Tudásbeli hiányosságok
A BIEN adatbázis kifejezetten arra szolgál, hogy a növényi biodiverzitással kapcsolatos tudásunkban meglévő hiányosságokat pótolja, mégis, ahogy Hortal et al. (2015) rámutat, az egymásra ható tudásbeli hiányosságok a legnagyobb léptékű biodiverzitás számszerűsítésében bizonytalansághoz vezetnek. Például nemcsak, hogy a feltérképezett fajaink kevesebb mint 10%-a rendelkezett mind tulajdonságadatokkal, mind filogenetikai információkkal, de ezek a hiányzó tulajdonságadatok (a fajok tulajdonságértékeinek ~66%-a) mind filogenetikai, mind földrajzi szempontból meglehetősen strukturáltak voltak. Így a funkcionális tulajdonságadatok Raunkiaeri hiányossága (sensu Hortal et al., 2015) kölcsönhatásba lép az elterjedési információk Wallace-i hiányosságával és a filogenetikai megértés darwini hiányosságával. És bár a filogenetikai ismereteket felhasználhatjuk e hiányzó értékek ésszerű imputálására (Swenson et al., 2017), az így kapott mintázatok számos, a biomok osztályozása szempontjából fontos aspektusa bizonytalan marad. Például a meglévő növekedési formaadatok azt sugallják, hogy a fás fajok dominálnak minden trópusi biomban, míg a lágyszárú növekedési formák arányos diverzitása sokkal nagyobb a mérsékelt égövi és a sarkvidéki biomokban (lásd még Engemann et al., 2016; Zanne et al., 2018). Vajon a lágyszárú növények relatív diverzitásának ez a szélességbeli növekedése a mintavételi torzulásokat és a trópusi lágyszárú növényekre vonatkozó taxonómiai ismeretek hiányát tükrözi, vagy pedig a nearktikus és neotrópusi birodalmak eltérő evolúciós és biogeográfiai történetét? A domináns növekedési formák a biomok egyik legfontosabb megkülönböztető jellemzője, így a legváltozatosabb (akár domináns, akár nem domináns) növekedési formákban bekövetkező szisztematikus változások szükségszerűen befolyásolni fogják a funkcionális diverzitás biomokon belüli változására vonatkozó előrejelzéseinket, és azt, hogy ezek a változások hogyan befolyásolják az ökoszisztéma működését és szolgáltatásait.
Főként azért használtunk fajterjedelmi térképeket bemeneti adatként, hogy leküzdjük a több forrásból összeállított adathalmazokra jellemző mintavételi torzítás tagadhatatlan problémáját (lásd az 1. kiegészítő ábrát). A fajok elterjedési modelljeiből készített elterjedési térképek azonban további bizonytalansági elemet jelenthetnek eredményeinkben. Például, tekintettel e vizsgálat földrajzi felbontására és egyes biomok térbeli összetettségére, ezek a modellek túlbecsülhették egyes fajok földrajzi kiterjedését egy-egy biom celláiból a közeli és egymáshoz kapcsolódó biomok celláiba. Az előfordulási adatok használata nem változtatta meg a tanulmány általános eredményeit (lásd a 2-6. kiegészítő ábrát); azonban a biomok közötti funkcionális átfedés kismértékű csökkenése valóban felfedhette néhány faj elterjedési területének túlbecslését (lásd a 2., 5. és 6. kiegészítő ábrát). Az előfordulási információk használata megváltoztatta a funkcionális megkülönböztethetőség és elterjedtség közötti általános összefüggéseket is az eredményeinkben, a mérsékelt és a trópusi régiók relatív túlsúlya miatt (lásd a 4. kiegészítő ábrát). Ebben az esetben a mi elterjedtségi mérőszámunkat korlátozza a trópusi bioszférák legtöbb fajának rendkívül korlátozott mintavétele. Az adatsorok és a mintavételezés javulásával a jövőben valószínűleg lehetőség nyílik e bizonytalanságok csökkentésére.
A másik adathiány, amelyet a biomok funkcionális sokféleségének megértéséhez kezelni kell, a gyakorisági információk prestoni hiányossága. A bőség különösen fontos a funkcionális sokféleség vizsgálatában, mivel a közösségek összeállása, az ökoszisztéma működése és a biogeokémiai körforgás szempontjából a legdominánsabb fajok tulajdonságai a legfontosabbak. Mivel összeállításainkat a fajeloszlási modellekből származtatott elterjedési térképekre alapoztuk, csak a gyakoriság földrajzi komponensével tudtunk foglalkozni, de a helyi abundancia és a földrajzi elterjedés közötti pozitív kapcsolat, az úgynevezett foglaltság-abundancia kapcsolat arra utal, hogy az egyes biomokban a legelterjedtebb fajok gyakran a gyakoribbak közé is tartozhatnak (Borregaard és Rahbek, 2010; Novosolov et al., 2017). Mivel a biomokat általában a régióban domináns növekedési formák jellemzik, a abundanciainformációk integrálása azt eredményezheti, hogy a biomok kevesebb funkcionális átfedést mutatnak. Azonban annak ellenére, hogy a BIEN olyan terjedelmes parcellaadatokat állított össze, amelyek lehetővé teszik a helyi abundancia becslését, a különböző régiók és növekedési formák közötti ésszerű összehasonlításokat az egyenlőtlen mintavételezés, a gamma-diverzitás regionális különbségei és a abundancia számszerűsítésének összehasonlíthatatlan módszerei nehezítik.
A biomokon belüli közös minták
A rendelkezésre álló adatok tartós hiányosságai ellenére elemzéseink feltártak egy korábban nem dokumentált összefüggést, amely a nyugati félteke valamennyi biomjára jellemző: bármely biomban a legelterjedtebb fajok általában alacsony funkcionális megkülönböztethetőséget is mutatnak, míg a funkcionálisan legjellemzőbb fajok szinte kivétel nélkül korlátozott elterjedésűek (3. ábra). Ez a “foglaltság-redundancia” összefüggés arra utalhat, hogy az egy biomban uralkodó éghajlati viszonyok egy sor közös jellemzőt választanak ki, és a funkcionálisan jellegzetesebb fajok az élőhelyek egy ritkább részhalmazára korlátozódnak az adott biomon belül. A funkcionálisan hasonló fajok általános elterjedtsége minden biomban és a “foglaltság-redundancia” kapcsolat egyaránt összhangban van a közösségi szintű funkcionális diverzitás közelmúltbeli globális elemzésével, amely szerint az élőhelyek szűrése funkcionálisan hasonló fajok együttéléséhez vezet (Li et al., 2018), valamint azokkal a tanulmányokkal, amelyek szerint a funkcionális redundancia növeli a közösség stabilitását (pl. Walker et al., 1999; Pillar et al., 2013). Ráadásul a biomok közötti nagyfokú taxonómiai és funkcionális átfedésekkel együtt (1., 5. ábra) az a tény, hogy a gyakori, széles körben elterjedt fajok funkcionálisan hasonlóak, megerősíti azt az elképzelést, hogy a szárazföldi növények a környezeti feltételek széles skáláján a funkcionális tulajdonságspektrum magja közelében közös jellemzőkkel rendelkeznek (Wright et al., 2004; Díaz et al., 2016). Umaña et al. (2017) eredményeitől eltérően a mi eredményeink azt mutatják, hogy a ritka, földrajzilag korlátozott fajok funkcionálisan különbözhetnek vagy nem különbözhetnek a szélesebb körben elterjedt és gyakori fajoktól. Így nyitva marad a kérdés, hogy a funkcionálisan megkülönböztetőbb fajok specialisták-e bizonyos környezetekben, vagy a tulajdonságkombinációik demográfiai vagy fiziológiai kompromisszumokat eredményeznek, amelyek korlátozzák földrajzi elterjedésüket.
Bioomok közötti összehasonlítások
A tulajdonságok hipervolumen eloszlásának összehasonlítása a biomok között (2B ábra) összhangban van azzal a megfigyeléssel, hogy a fajgazdagabb környezetek funkcionálisan is változatosabbak (Swenson et al., 2012; Lamanna et al., 2014; Li et al., 2018; Šímová et al., 2018). Eredményeink kétértelműbbek azzal a hipotézissel kapcsolatban, hogy a szezonális és szélsőséges éghajlati környezetek következetesen korlátozzák a fajok funkcionális diverzitását (de la Riva et al., 2018). Az összes trópusi biom magasabb átlagos funkcionális diverzitást mutat, mint az összes mérsékelt égövi biom, és a sarkvidéki biomok valóban a legkisebb hipervolumeneket mutatják. Az egyes csoportokon belül azonban a szárazabb vagy szezonálisan változékonyabb biomok nem mindig mutatnak kisebb hipervolument (pl. száraz erdők, xerikus erdők), ahogy azt Currie et al. (2004) toleranciahipotézise alapján vártuk volna, míg az optimális éghajlati viszonyok a fiziológiai paraméterek több kombinációját támogatják. Egyedül ezekből az eredményekből nem tudjuk eldönteni, hogy a mérsékelt égövi és a sarkvidéki biomok a funkcionális niche-tér korlátai miatt kevésbé taxonómiailag változatosak, vagy pedig funkcionális hipervolumeneik azért kicsik, mert taxonómiailag nem változatosak. A null-modellezési megközelítések potenciálisan segíthetnek a taxonómiai és funkcionális diverzitás szétválasztásában (Swenson et al., 2012; Lamanna et al., 2014; Šímová et al., 2018), de egy ilyen elemzés meghaladta e tanulmány kereteit. Ennél is fontosabb, hogy eredményeink megerősítik annak megértésének fontosságát, hogy az evolúciós és biogeográfiai történelem hogyan alakítja a biomok funkcionális diverzitását (Woodward et al., 2004; Pennington et al., 2006; Donoghue és Edwards, 2014; Moncrieff et al., 2016). A fajok (és magasabb vonalvezetések) kiterjedt megosztása a biomokon belül, de nagyrészt nem a különböző biogeográfiai birodalmak között, egyrészt a birodalmakon belüli funkcionális sokféleség homogenizálására szolgálhat, másrészt nyomokat szolgáltathat a különböző környezetek által szelektált vagy ellenük szelektált jellegzetes vonásokról (Douma et al., 2012; Zanne et al., 2014, 2018).
Az összes biom közötti jelentős hipervolumen átfedés ellenére (2. kiegészítő ábra) úgy tűnik, hogy a trópusi, mérsékelt égövi és hideg biomok mind a funkcionális tér elkülöníthető régióit foglalják el (5B,C ábra). Úgy tűnik, hogy a biomokat megkülönböztető fő vonások a növények teljes méretével kapcsolatos vonások, beleértve mind az érett magasságot, mind a magok tömegét (7-10. kiegészítő ábra), és nem a levélgazdaságossági vonások, ahogyan azt a lokálisabb, parcellákon alapuló elemzésekben megfigyelték (pl. Douma et al., 2012). Kivételt képez a levél P, amely jelentős különbségeket mutatott a trópusi és a mérsékelt égövi/poláris biomok között (7-10. kiegészítő ábra), amely mintázatot más, fajspecifikus értékeken és egész ökoszisztéma méréseken alapuló elemzésekben is megfigyeltünk (Kerkhoff et al., 2005; Swenson et al., 2012). Mivel a levél P a legritkábban mintavételezett tulajdonságunk volt, a trópusi és mérsékelt égövi/poláris területek közötti különbségek az imputációs eljárásból eredő torzulásoknak lehetnek kitéve. A levél P azonban jelentős filogenetikai jelet mutat (Kerkhoff et al., 2006), ami arra utal, hogy az imputációknak torzításmentesnek kell lenniük (Swenson et al., 2017). A Leaf P szintén szignifikánsan magasabb a lágyszárú növekedési formákban (Kerkhoff et al., 2006), így a szélességi eltolódás a túlnyomórészt fás és lágyszárú diverzitás között (4. ábra) szintén befolyásolhatja ezt a mintázatot.
Amikor a hipervolumen-elemzéseket csak az egyes bioszférákban legelterjedtebb és funkcionálisan leggyakoribb fajokra korlátoztuk, az egyes biogeográfiai bioszférákon belüli egyes bioszférák kevésbé fedték egymást a funkcionális tulajdonságok terében (vö. 5B,C ábrák), ami arra utal, hogy ezek a fajok az egyes környezetekhez jobban alkalmazkodott fenotípusokat tükrözhetnek. Ebben az összefüggésben a xerikus erdei bioszféra különösen érdekes. Részben a Chaco ökorégió bevonása miatt a xerikus erdőterületek taxonómiai szempontból a trópusi biomok közé csoportosulnak (5A. ábra). Ha azonban az összes faj funkcionális hipervolumenét figyelembe vesszük, sokkal erősebb hasonlóságot mutatnak a mérsékelt égövi biomokkal (5B. ábra). Végül, amikor csak a legelterjedtebb és funkcionálisan leggyakoribb fajokat elemezzük, a xerikus erdők ismét fokozott hasonlóságot mutatnak a trópusi biomokkal, de a mérsékelt égövi gyepekkel és a mediterrán biomokkal is nagyfokú hasonlóságot mutatnak (5C. ábra). A meleg, mezikus környezetekből a hidegebb, szárazabb és szezonálisabb környezetekbe való átmenetet hasonló tulajdonságok, például a lágyszárúak habitusa segíti elő (Douma et al., 2012; Zanne et al., 2014, 2018), és a hidegebb környezetekhez hasonlóan a száraz környezetek közösségei is inkább filogenetikai szempontból klaszteresednek (Qian és Sandel, 2017). Ráadásul a xerikus zónák elhelyezkedése a trópusok közötti konvergenciazóna határán földrajzi átmenetet képeznek a trópusi és a mérsékelt égöv között. Az a tény, hogy a xerikus erdőségek mind funkcionális, mind biogeográfiai szempontból köztes helyet foglalnak el a trópusi és mérsékelt égövi birodalmak között, tovább erősíti azt a gondolatot, hogy a biomok funkcionális sokféleségének jobb megértéséhez figyelembe kell vennünk biogeográfiai és filogenetikai történetüket (Pennington et al., 2006; Donoghue és Edwards, 2014; Moncrieff et al, 2016).
Következtetések
A szárazföldi biomok bármilyen osztályozása kis számú diszkrét kategóriát kényszerít az éghajlat és a fajok elterjedésének folyamatos gradiensére, és így a komplex ökológiai táj durva egyszerűsítését jelenti (Moncrieff et al., 2016, de lásd Silva de Miranda et al., 2018). A túlzott leegyszerűsítés lehetősége ellenére azonban a biomok hasznos konstrukciók a Föld főbb szárazföldi ökoszisztémáinak biodiverzitásának és működésének rendszerezéséhez és megértéséhez, és a tulajdonságalapú megközelítések nagy potenciállal rendelkeznek a globális vegetációs eloszlások dinamikus modellezésének segítésére.
Ebben a tanulmányban kimutattuk, hogy valamennyi biom taxonómiai változatosságának nagy része olyan fajokat képvisel, amelyek mind szűken elterjedtek, mind funkcionálisan hasonlóak. Továbbá, az egyes biomokon belül a funkcionálisan legjellegzetesebb fajok az egyes biomokban földrajzilag is általában ritkák, míg a széles körben elterjedt fajok egységesen alacsony funkcionális különállóságot mutatnak. A biomok közötti kiterjedt taxonómiai és funkcionális átfedések ellenére a biomok megkülönböztethető, biogeográfiailag és klimatikusan elkülönülő egységekbe csoportosulnak, különösen akkor, ha a funkcionális klaszterezést az egyes biomokban legelterjedtebb és funkcionálisan leggyakoribb fajokra alapozzuk. A biomok funkcionális megértésének előmozdításához azonban nemcsak a vegetációtípusokon belüli és a vegetációtípusok közötti tulajdonságvariáció jobb jellemzésére lesz szükség, hanem a fajegyüttesek biogeográfiai és filogenetikai történetére és a fajok biomokon belüli relatív gyakoriságára vonatkozó információkra is.
Adatok elérhetősége
A jelen tanulmányhoz elemzett tulajdonságadatkészletek és fajterjedelmi térképek letölthetők a BIEN R csomagon keresztül (Maitner et al., 2018). A BIEN adatbázissal kapcsolatos további részletekért lásd http://bien.nceas.ucsb.edu/bien/.
A szerzők hozzájárulása
SE-L, AK, BJE, DMN és CV tervezték a tanulmányt; SE-L és AK elemezték az adatokat; SE-L és AK írták a kézirat első változatát; CV, BB, NK, BSM, BM, RKP, BS, SS, J-CS és SKW hozzájárultak a tulajdonságok és a hatótávolsági térképek adatbázisának összeállításához és létrehozásához. Minden szerző hozzájárult a kézirat kidolgozásához és megírásához.
Finanszírozás
SE-L, DMN és AK az Egyesült Államok Nemzeti Tudományos Alapítványának (DEB-1556651) közös kutatási ösztöndíjával támogatott. BJE-t a Nemzeti Tudományos Alapítvány DEB-1457812 és Macrosystems-1065861 díja támogatta. CV-t az Európai Kutatási Tanács (Grant ERC-StG-2014-639706-CONSTRAINTS) és a Francia Biodiverzitás Kutatási Alapítvány (FRB; www.fondationbiodiversite.fr) támogatta a CESAB projekt keretében Causes and consequences of functional rarity from local to global scales. J-CS ezt a munkát a VILLUM FONDEN által finanszírozott VILLUM Investigator Biodiversity Dynamics in a Changing World (16549-es támogatás) projektjéhez való hozzájárulásnak tekinti. SKW-t az Új-Zélandi Üzleti, Innovációs és Foglalkoztatási Minisztérium Tudományos és Innovációs Csoportjának Stratégiai Tudományos Befektetési Alapja támogatta.
Kérdések összeférhetetlenségi nyilatkozata
A szerzők kijelentik, hogy a kutatást olyan kereskedelmi vagy pénzügyi kapcsolatok hiányában végezték, amelyek potenciális összeférhetetlenségként értelmezhetők.
Köszönet
Köszönjük Dan Griffithnek és két lektornak az építő jellegű kritikát, amely nagyban javította ezt a kéziratot. Köszönetet mondunk továbbá minden BIEN adatközlőnek (a teljes listát lásd http://bien.nceas.ucsb.edu/bien/data-contributors/).
Kiegészítő anyagok
A cikkhez tartozó kiegészítő anyagok online elérhetők a következő címen: https://www.frontiersin.org/articles/10.3389/fevo.2018.00219/full#supplementary-material
Banda, R.K., Delgado-Salinas, A., Dexter, K. G., Linares-Palomino, R., Oliveira-Filho, A., Prado, D., et al. (2016). A neotrópusi száraz erdők növényi diverzitási mintázatai és természetvédelmi vonatkozásaik. Science 353, 1383-1387. doi: 10.1126/science.aaf5080
PubMed Abstract | CrossRef Full Text | Google Scholar
Blonder, B., Lamanna, C., Violle, C., and Enquist, B. J. (2014). Az n-dimenziós hipertérfogat. Glob. Ecol. Biogeogr. 23, 595-609. doi: 10.1111/geb.12146
CrossRef Full Text | Google Scholar
Blonder, B., Morrow, C. B., Maitner, B., Harris, D. J., Lamanna, C., Violle, C., et al. (2018). Új megközelítések n-dimenziós hipertérfogatok lehatárolására. Methods Ecol. Evol. 9, 305-319. doi: 10.1111/2041-210X.12865
CrossRef Full Text | Google Scholar
Bonan, G. B., Oleson, K. W., Fisher, R. A., Lasslop, G., and Reichstein, M. (2012). A levélfiziológiai jellemzők és a lombkorona fluxusadatok egyeztetése: a TRY és FLUXNET adatbázisok használata a Community Land Model 4. verziójában. J. Geophys. Res. Biogeosci. 117:G02026. doi: 10.1029/2011JG001913
CrossRef Full Text | Google Scholar
Borregaard, M. K., and Rahbek, C. (2010). A földrajzi elterjedés és a fajszám közötti kapcsolat kauzalitása. Q. Rev. Biol. 85, 3-25. doi: 10.1086/650265
PubMed Abstract | CrossRef Full Text | Google Scholar
Bruggeman, J., Heringa, J., and Brandt, B. W. (2009). PhyloPars: hiányzó paraméterértékek becslése filogenitás segítségével. Nucleic Acids Res. 37, W179-W184. doi: 10.1093/nar/gkp370
PubMed Abstract | CrossRef Full Text | Google Scholar
Cadotte, M. W., Carscadden, K., and Mirotchnick, N. (2011). A fajokon túl: funkcionális sokféleség és az ökológiai folyamatok és szolgáltatások fenntartása. J. Appl. Ecol. 48, 1079-1087. doi: 10.1111/j.1365-2664.2011.02048.x
CrossRef Full Text | Google Scholar
Cornwell, W. K., Cornelissen, J. H., Amatangelo, K., Dorrepaal, E., Eviner, V. T., Godoy, O., et al. (2008). A növényfajok jellemzői az alom lebomlási sebességének domináns szabályozói a biomokon belül világszerte. Ecol. Lett. 11, 1065-1071. doi: 10.1111/j.1461-0248.2008.01219.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Craine, J. M. (2009). A vadon élő növények erőforrás-stratégiái. New Jersey, NJ: Princeton University Press. doi: 10.1515/9781400830640
CrossRef Full Text | Google Scholar
Currie, D. J., Mittelbach, G. G., Cornell, H. V., Field, R., Guégan, J. F., Hawkins, B. A., et al. (2004). A taxonómiai gazdagság széles skálájú változásának éghajlati alapú hipotéziseinek előrejelzése és tesztelése. Ecol. Lett. 12, 1121-1134. doi: 10.1111/j.1461-0248.2004.00671.x
CrossRef Full Text | Google Scholar
de la Riva, E. G., Violle, C., Pérez-Ramos, I. M., Marañón, T., Navarro-Fernández, C. M., Olmo, M., et al. (2018). A többdimenziós funkcionális tulajdonságok megközelítése feltárja a környezeti stressz lenyomatát a mediterrán fásszárú közösségekben. Ecosystems 21, 248-262. doi: 10.1007/s10021-017-0147-7
CrossRef Full Text | Google Scholar
Díaz, S., Kattge, J., Cornelissen, J. H., Wright, I. J., Lavorel, S., Dray, S., et al. (2016). A növényi forma és funkció globális spektruma. Nature 529, 167-171. doi: 10.1038/nature16489
PubMed Abstract | CrossRef Full Text | Google Scholar
Díaz, S., Lavorel, S., de Bello, F., Quétier, F., Grigulis, K., and Robson, T. M. (2007). A növényi funkcionális sokféleség hatásainak beépítése az ökoszisztéma-szolgáltatások értékelésébe. Proc. Natl. Acad. Sci. U.S.A 104, 20684-20689. doi: 10.1073/pnas.0704716104
PubMed Abstract | CrossRef Full Text | Google Scholar
Donoghue, M. J., and Edwards, E. J. (2014). Biomváltások és niche-evolúció a növényekben. Annu. Rev. Ecol. Evol. Syst. 45, 547-572. doi: 10.1146/annurev-ecolsys-120213-091905
CrossRef Full Text | Google Scholar
Douma, J. C., Aerts, R., Witte, J. P. M., Bekker, R. M., Kunzmann, D., Metselaar, K., et al. (2012). A funkcionálisan különböző növényi jellemzők kombinációja lehetővé teszi a fajösszetételek széles körének kvantitatív előrejelzését Északnyugat-Európában. Ecography 35, 364-373. doi: 10.1111/j.1600-0587.2011.07068.x
CrossRef Full Text | Google Scholar
Engemann, K., Sandel, B., Enquist, B. J., Jørgensen, P. M., Kraft, N., Marcuse-Kubitza, A., et al. (2016). A növényi funkcionális csoportok dominanciájának mintázatai és mozgatórugói a nyugati féltekén: egy hatalmas botanikai adathalmazon alapuló makroökológiai újraértékelés. Bot. J. Linn. Soc. 180, 141-160. doi: 10.1111/boj.12362
CrossRef Full Text | Google Scholar
Enquist, B. J., Condit, R., Peet, R., Thiers, B., and Schildhauer, M. (2016). A botanikai információs és ökológiai hálózat (BIEN): Kiberinfrastruktúra egy integrált botanikai információs hálózathoz a globális éghajlatváltozásnak a növényi biodiverzitásra gyakorolt ökológiai hatásainak vizsgálatára. PeerJ Prepr. 4:e2615v2. doi: 10.7287/peerj.preprints.2615v2
CrossRef Full Text | Google Scholar
Garnier, E., Cortez, J., Billès, G., Navas, M. L., Roumet, C., Debussche, M., et al. (2004). Növényi funkcionális markerek rögzítik az ökoszisztéma tulajdonságait a másodlagos szukcesszió során. Ecology 85, 2630-2637. doi: 10.1890/03-0799
CrossRef Full Text | Google Scholar
Goldsmith, G. R., Morueta-Holme, N., Sandel, B., Fitz, E. D., Fitz, S. D., Boyle, B., et al. (2016). Plant-O-Matic: az amerikai kontinens összes növényének dinamikus és mobil útmutatója. Methods Ecol. Evol. 7, 960-965. doi: 10.1111/2041-210X.12548
CrossRef Full Text | Google Scholar
Goolsby, E. W., Bruggeman, J., and Ané, C. (2017). Rphylopars: gyors többváltozós filogenetikai összehasonlító módszerek hiányzó adatok és fajon belüli variáció esetén. Methods Ecol. Evol. 8, 22-27. doi: 10.1111/2041-210X.12612
CrossRef Full Text | Google Scholar
Grenié, M., Denelle, P., Tucker, C. M., Munoz, F., and Violle, C. (2017). funrar: an R package to characterize functional rarity. Divers. Distrib. 23, 1365-1371. doi: 10.1111/ddi.12629
CrossRef Full Text | Google Scholar
Hortal, J., de Bello, F., Diniz-Filho, J. A. F., Lewinsohn, T. M., Lobo, J. M., and Ladle, R. J. (2015). A biológiai sokféleséggel kapcsolatos nagyléptékű ismereteket sújtó hét hiányosság. Annu. Rev. Ecol. Evol. Syst. 46, 523-549. doi: 10.1146/annurev-ecolsys-112414-054400
CrossRef Full Text | Google Scholar
Kerkhoff, A. J., Enquist, B. J., Elser, J. J., and Fagan, W. F. (2005). Növényi allometria, sztöchiometria és az elsődleges termelékenység hőmérsékletfüggése. Glob. Ecol. Biogeogr. 14, 585-598. doi: 10.1111/j.1466-822X.2005.00187.x
CrossRef Full Text | Google Scholar
Kerkhoff, A. J., Fagan, W. F., Elser, J. J., and Enquist, B. J. (2006). Filogenetikai és növekedési formavariációk a nitrogén és foszfor skálázásában a magoncnövényekben. Am. Nat. 168, E103-E122. doi: 10.1086/507879
PubMed Abstract | CrossRef Full Text | Google Scholar
Kraft, N. J., Valencia, R., and Ackerly, D. D. (2008). Funkcionális tulajdonságok és niche-alapú faközösség-összeállítás egy amazóniai erdőben. Science 322, 580-582. doi: 10.1126/science.1160662
PubMed Abstract | CrossRef Full Text | Google Scholar
Lamanna, C., Blonder, B., Violle, C., Kraft, N. J., Sandel, B., et al. (2014). Funkcionális tulajdonságtér és a szélességi diverzitás gradiens. Proc. Natl. Acad. Sci. U.S.A. 111, 13745-13750. doi: 10.1073/pnas.1317722111
PubMed Abstract | CrossRef Full Text | Google Scholar
Lavorel, S., and Garnier, E. (2002). A közösségek összetételében és az ökoszisztémák működésében bekövetkező változások előrejelzése növényi jellemzőkből: a Szent Grál újragondolása. Funct. Ecol. 16, 545-556. doi: 10.1046/j.1365-2435.2002.00664.x
CrossRef Full Text | Google Scholar
Lavorel, S., Grigulis, K., Lamarque, P., Colace, M.-P., Garden, D., Girel, J., et al. (2011). Növényi funkcionális jellemzők felhasználása a több ökoszisztéma-szolgáltatás táji eloszlásának megértéséhez. J. Ecol. 99, 135-147. doi: 10.1111/j.1365-2745.2010.01753.x
CrossRef Full Text | Google Scholar
Li, Y., Shipley, B., Price, J. N., Dantas, V. de L., Tamme, R., Westoby, M., et al. (2018). Az élőhelyszűrés határozza meg a növényközösségek funkcionális niche-foglaltságát világszerte. J. Ecol. 106, 1001-1009. doi: 10.1111/1365-2745.12802
CrossRef Full Text | Google Scholar
Magallón, S., Gómez-Acevedo, S., Sánchez-Reyes, L. L., and Hernández-Hernández, T. (2015). Egy metakalibrált időfa dokumentálja a virágos növények filogenetikai diverzitásának korai emelkedését. New Phytol. 207, 437-453. doi: 10.1111/nph.13264
PubMed Abstract | CrossRef Full Text | Google Scholar
Maitner, B. S., Boyle, B., Casler, N., Condit, R., Donoghue, J., Durán, S. M., et al. (2018). A BIEN R csomag: eszköz a botanikai információs és ökológiai hálózat (BIEN) adatbázisának eléréséhez. Methods Ecol. Evol. 9, 373-379. doi: 10.1111/2041-210X.12861
CrossRef Full Text | Google Scholar
Moncrieff, G. R., Bond, W. J., and Higgins, S. I. (2016). A biom fogalmának felülvizsgálata a globális változások hatásainak megértéséhez és előrejelzéséhez. J. Biogeogr. 43, 863-873. doi: 10.1111/jbi.12701
CrossRef Full Text | Google Scholar
Novosolov, M., Rodda, G. H., North, A. C., Butchart, S. H. M., Tallowin, O. J. S., Gainsbury, A. M., et al. (2017). Populációs sűrűség-tartományméret kapcsolat felülvizsgálva. Glob. Ecol. Biogeogr. 26, 1088-1097. doi: 10.1111/geb.12617
CrossRef Full Text | Google Scholar
Olson, D. M., Dinerstein, E., Wikramanayake, E. D., Burgess, N. D., Powell, G. V. N., Underwood, E. C., et al. (2001). Terrestrial Ecoregions of the world: a földi élet új térképe: a szárazföldi ökorégiók új globális térképe innovatív eszköz a biológiai sokféleség megőrzéséhez. Bioscience 51, 933-938. doi: 10.1641/0006-3568(2001)0512.0.CO;2
CrossRef Full Text | Google Scholar
Pennington, R. T., Prado, D. E., and Pendry, C. A. (2000). Neotropical seasonally dry forests and quaternary vegetation changes. J. Biogeogr. 27, 261-273. doi: 10.1046/j.1365-2699.2000.00397.x
CrossRef Full Text | Google Scholar
Pennington, R. T., Richardson, J. E., and Lavin, M. (2006). Betekintés a fajgazdag biomok történeti felépítésébe a datált növényi filogeniák, a semleges ökológiai elmélet és a filogenetikai közösségszerkezet alapján. New Phytol. 172, 605-616. doi: 10.1111/j.1469-8137.2006.01902.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Penone, C., Davidson, A. D., Shoemaker, K. T., Di Marco, M., Rondinini, C., Brooks, T. M., et al. (2014). Hiányzó adatok imputálása élettörténeti tulajdonságokra vonatkozó adatsorokban: melyik megközelítés teljesít a legjobban? Methods Ecol. Evol. 5, 961-970. doi: 10.1111/2041-210X.12232
CrossRef Full Text | Google Scholar
Pillar, V. D., Blanco, C. C., Müller, S. C., Sosinski, E. E., Joner, F., and Duarte, L. D. S. (2013). Funkcionális redundancia és stabilitás a növényi közösségekben. J. Veg. Sci. 24, 963-974. doi: 10.1111/jvs.12047
CrossRef Full Text | Google Scholar
Prado, D. E., and Gibbs, P. E. (1993). A fajok elterjedési mintázatai Dél-Amerika száraz évszakos erdeiben. Ann. Mo. Bot. Gard. 80, 902-927. doi: 10.2307/2399937
CrossRef Full Text | Google Scholar
Qian, H., and Sandel, B. (2017). Regionális angiospermia-összetételek filogenetikai szerkezete Észak-Amerika szélességi és éghajlati gradiensein keresztül. Glob. Ecol. Biogeogr. 26, 1258-1269. doi: 10.1111/geb.12634
CrossRef Full Text | Google Scholar
Reich, P. B., Wright, I. J., Cavender-Bares, J., Craine, J. M., Oleksyn, J., Westoby, M., et al. (2003). A növényi funkcionális variáció evolúciója: tulajdonságok, spektrumok és stratégiák. Int. J. Plant Sci. 164, S143-S164. doi: 10.1086/374368
CrossRef Full Text | Google Scholar
Silva de Miranda, P. L., Oliveira-Filho, A., Pennington, R. T., Neves, D. M., Baker, T. R., and Dexter, K. G. (2018). A fafajleltárak felhasználása a biomok feltérképezésére és éghajlati átfedéseik értékelésére az alföldi trópusi Dél-Amerikában. Glob. Ecol. Biogeogr. 27, 899-912. doi: 10.1111/geb.12749
CrossRef Full Text | Google Scholar
Šímová, I., Violle, C., Kraft, N. J. B., Storch, D., Svenning, J.-C., Boyle, B., et al. (2015). A tulajdonságok átlagainak és varianciáinak eltolódása észak-amerikai faegyüttesekben: a fajgazdagság mintázatai lazán kapcsolódnak a funkcionális térhez. Ecography 38, 649-658. doi: 10.1111/ecog.00867
CrossRef Full Text | Google Scholar
Šímová, I., Violle, C., Svenning, J.-C., Kattge, J., Engemann, K., Sandel, B., et al. (2018). A főbb növényi jellemzők térbeli mintázatai és éghajlati kapcsolatai az Újvilágban eltérnek a fás és lágyszárú fajok között. J. Biogeogr. 45, 895-916. doi: 10.1111/jbi.13171
CrossRef Full Text | Google Scholar
Smith, S. A., and Beaulieu, J. M. (2009). Az élettörténet befolyásolja a klimatikus niche evolúciójának sebességét a virágos növényekben. Proc. Biol. Sci. 276, 4345-4352. doi: 10.1098/rspb.2009.1176
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, S. A., and Brown, J. W. (2018). Egy széleskörűen inkluzív magoncnövény-filogenetika konstruálása. Am. J. Bot. 105, 302-314. doi: 10.1002/ajb2.1019
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, S. A., and Donoghue, M. J. (2008). A molekuláris evolúció sebessége a virágos növények élettörténetéhez kapcsolódik. Science 322, 86-89. doi: 10.1126/science.1163197
PubMed Abstract | CrossRef Full Text | Google Scholar
Swenson, N. G. (2014). Phylogenetic imputation of plant functional trait databases. ecography 37, 105-110. doi: 10.1111/j.1600-0587.2013.00528.x
CrossRef Full Text
Swenson, N. G., Enquist, B. J., Pither, J., Kerkhoff, A. J., Boyle, B., Weiser, M. D., et al. (2012). A fásszárú növények funkcionális diverzitásának biogeográfiája és szűrése Észak- és Dél-Amerikában. Glob. Ecol. Biogeogr. 21, 798-808. doi: 10.1111/j.1466-8238.2011.00727.x
CrossRef Full Text | Google Scholar
Swenson, N. G., Weiser, M. D., Mao, L., Araújo, M. B., Diniz-Filho, J. A. F., Kollmann, J., et al. (2017). A filogenetika és a fa funkcionális diverzitásának előrejelzése új kontinentális környezetben. Glob. Ecol. Biogeogr. 26, 553-562. doi: 10.1111/geb.12559
CrossRef Full Text | Google Scholar
Umaña, M. N., Mi, X., Cao, M., Enquist, B. J., Hao, Z., Howe, R., et al. (2017). A funkcionális egyediség és a térbeli aggregáció szerepe a fák ritkaságának magyarázatában. Glob. Ecol. Biogeogr. 26, 777-786. doi: 10.1111/geb.12583
CrossRef Full Text | Google Scholar
van Bodegom, P. M., Douma, J. C., and Verheijen, L. M. (2014). Teljesen vonásalapú megközelítés a globális vegetációs eloszlás modellezésére. Proc. Natl. Acad. Sci. U.S.A. 111, 13733-13738. doi: 10.1073/pnas.1304551110
PubMed Abstract | CrossRef Full Text | Google Scholar
Violle, C., Reich, P. B., Pacala, S. W., Enquist, B. J., and Kattge, J. (2014). A funkcionális biogeográfia kialakulása és ígérete. Proc. Natl. Acad. Sci. U.S.A. 111, 13690-13696. doi: 10.1073/pnas.1415442111
PubMed Abstract | CrossRef Full Text | Google Scholar
Violle, C., Thuiller, W., Mouquet, N., Munoz, F., Kraft, N. J. B., Cadotte, M. W., et al. (2017). Funkcionális ritkaság: a kiugró fajok ökológiája. Trends Ecol. Evol. 32, 356-367. doi: 10.1016/j.tree.2017.02.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Walker, B., Kinzig, A., and Langridge, J. (1999). Növényi attribútumdiverzitás, rugalmasság és ökoszisztémafunkció: a domináns és mellékfajok természete és jelentősége. Ecosystems 2, 95-113. doi: 10.1007/s100219900062
CrossRef Full Text | Google Scholar
Westoby, M. (1998). A levélmagasság-magasság (LHS) növényökológiai stratégiai rendszer. Plant Soil 199, 213-227. doi: 10.1023/A:1004327224729
CrossRef Full Text | Google Scholar
Woodward, F. I., Lomas, M. R., and Kelly, C. K. (2004). Globális éghajlat és a növényi biomok eloszlása. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 359, 1465-1476. doi: 10.1098/rstb.2004.1525
PubMed Abstract | CrossRef Full Text | Google Scholar
Wright, I. J., Reich, P. B., Westoby, M., Ackerly, D. D., Baruch, Z., Bongers, F., et al. (2004). A világméretű levélgazdasági spektrum. Nature 428, 821-827. doi: 10.1038/nature02403
PubMed Abstract | CrossRef Full Text | Google Scholar
Xia, J., Niu, S., Ciais, P., Janssens, I. A., Chen, J., Ammann, C., et al. (2015). A szárazföldi bruttó primer termelékenység együttes szabályozása a növények fenológiája és fiziológiája által. Proc. Natl. Acad. Sci. U.S.A. 112, 2788-2793. doi: 10.1073/pnas.1413090112
PubMed Abstract | CrossRef Full Text | Google Scholar
Zanne, A. E., Pearse, W. D., Cornwell, W. K., McGlinn, D. J., Wright, I. J., and Uyeda, J. C. (2018). Az angiospermák funkcionális biogeográfiája: élet a szélsőségekben. New Phytol. 218, 1697-1709. doi: 10.1111/nph.15114
PubMed Abstract | CrossRef Full Text | Google Scholar
Zanne, A. E., Tank, D. C., Cornwell, W. K., Eastman, J. M., Smith, S. A., FitzJohn, R. G., et al. (2014). Három kulcs az angiospermák fagyos környezetbe sugárzásához. Nature 506, 89-92. doi: 10.1038/nature12872
PubMed Abstract | CrossRef Full Text | Google Scholar