Értékelés | Biopszichológia | Összehasonlító | Kognitív | Fejlődés | Nyelv | Egyéni különbségek | Személyiség | Filozófia | Szociális |
Módszerek | Statisztika | Klinikai | Oktatási | Oktatási | Ipari | Szakmai tételek | Világpszichológia |
Biológiai:Viselkedésgenetika – Evolúciós pszichológia – Neuroanatómia – Neurokémia – Neuroendokrinológia – Idegtudomány – Pszichoneuroimmunológia – Fiziológiai pszichológia – Pszichofarmakológia(Index, vázlat)
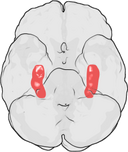
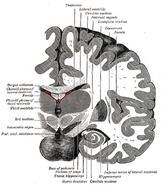
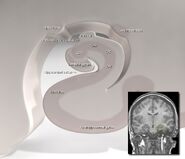
A hippokampusz elhelyezkedése az emberi agyban.
Az azonos nevű folyóirathoz lásd Hippocampus (folyóirat)
A hippocampus az előagy része, a medialis temporális lebenyben található. A limbikus rendszerhez tartozik, és fontos szerepet játszik a hosszú távú emlékezetben és a térbeli navigációban. Az embereknek és más emlősöknek két hippocampusa van, egy-egy az agy mindkét oldalán. A rágcsálóknál, ahol a legtöbbet tanulmányozták, a hippokampusz banán alakú. Az embernél görbült és tekervényes alakja a korai anatómusokat a csikóhalra emlékeztette. A neve valójában a görög csikóhal szóból származik (görögül: hippos = ló, campos = tenger).
Az Alzheimer-kórban a hippokampusz az egyik első olyan agyterület, amely károsodik; a memóriazavarok és a tájékozódási zavar az első tünetek között jelentkeznek. A hippokampusz károsodását okozhatja oxigénhiány (anoxia), agyvelőgyulladás vagy meziális temporális lebeny epilepszia is. A kiterjedt hippokampusz-károsodásban szenvedő embereknél amnézia, azaz az új emlékek kialakítására vagy megőrzésére való képtelenség jelentkezhet.
A hippokampusz funkciói
A legkorábbi elképzelés talán az volt, hogy a hippokampusz részt vesz a szaglásban: ezt, úgy tűnik, elsősorban az agyban, a szaglókéreg mellett való elhelyezkedése sugallta. A hippokampusz szaglási válaszai iránt továbbra is van némi érdeklődés, de ma már szinte senki sem hiszi, hogy a hippokampusz elsődleges funkciója a szaglás.
A hippokampusz működésével kapcsolatban az évek során három fő elképzelés uralta a szakirodalmat: a gátlás, az emlékezet és a tér. A viselkedési gátlás elmélete (amelyet O’Keefe és Nadel úgy karikírozott, hogy “lépj a fékre!”) az 1960-as évekig nagyon népszerű volt. Erejének nagy részét két megfigyelésből merítette: először is, a hippokampusz-károsodott állatok általában hiperaktívak; másodszor, a hippokampusz-károsodott állatok gyakran nehezen tanulják meg a korábban megtanult válaszok gátlását. Jeffrey Gray ezt a gondolatmenetet a hippokampusz szorongásban betöltött szerepének teljes értékű elméletévé fejlesztette tovább. A gátlási elmélet azonban jelenleg nem túl népszerű.
A második fontos gondolatmenet a hippokampuszt a memóriával hozza összefüggésbe. Bár voltak előzményei, ez az elképzelés fő erejét Scoville és Milner igen ismert beszámolójából merítette, amely a hippokampusz műtéti pusztításának eredményeiről szólt (az epilepsziás rohamok enyhítésére tett kísérlet során), egy H. M. nevű betegnél. A váratlan eredmény súlyos amnézia volt: H. M. képtelen volt tudatosan emlékezni a műtétje után vagy azt megelőző néhány évben történt eseményekre. Ez az eset olyan hatalmas érdeklődést váltott ki, hogy ma már azt mondják, hogy H.M. a történelem legintenzívebben tanulmányozott orvosi esete. Az ezt követő években más, hasonló mértékű hippokampusz-károsodással és (baleset vagy betegség okozta) amnéziával küzdő betegeket is tanulmányoztak, és szó szerint több ezer kísérletben vizsgálták a hippokampusz idegi plaszticitásának fiziológiáját. Ma már szinte általános egyetértés van abban, hogy a hippokampusz valamilyen fontos szerepet játszik a memóriában; ennek a szerepnek a pontos természete azonban továbbra is széles körben vitatott.
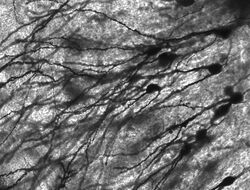
Golgi-festékes neuronok a rágcsáló hippokampuszában.
Szerep a memóriában
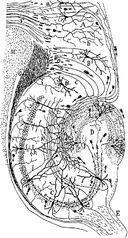
A rágcsáló hippokampuszának idegi áramköreinek rajza. S. Ramón y Cajal, 1911.
Pszichológusok és idegtudósok vitatják a hippokampusz pontos szerepét, de abban általában egyetértenek, hogy alapvető szerepe van a személyesen átélt eseményekről szóló új emlékek kialakításában (epizodikus vagy önéletrajzi emlékezet). Egyes kutatók inkább a hippokampuszt egy nagyobb mediális halántéklebenyi memóriarendszer részének tekintik, amely az általános deklaratív emlékezetért felelős (az explicit módon verbalizálható emlékek – ezek közé tartozna például az epizodikus emlékezet mellett a tényekre való emlékezés is).
Fő cikk: A hippokampusz szerepe az emlékezetben
Egyes bizonyítékok szerint a hippokampusz részt vesz a térbeli információk tárolásában és feldolgozásában. Patkányokon végzett vizsgálatok kimutatták, hogy a hippokampusz neuronjai térbeli tüzelési mezőkkel rendelkeznek. Ezeket a sejteket helysejteknek nevezik. Egyes sejtek akkor tüzelnek, amikor az állat egy adott helyen találja magát, függetlenül a mozgás irányától, míg a legtöbb sejt legalább részben érzékeny a fej irányára és a mozgás irányára.
Fő cikk: A hippokampusz szerepe a térbeli emlékezetben és navigációban
Hippokampusz és depresszió
- A hippokampusz térfogatának csökkenését,ami talán a túlzott glükokortikoidok
neurogenezisre gyakorolt hatását tükrözi, depressziós emberek csoportjaiban azonosították a kontrollcsoportokhoz képest.
Fő cikk: Depresszió és a hippokampusz
Történet
A hippokampusz kifejezést először Giulio Cesare Aranzi anatómus (1564 körül) használta az agyi szerv leírására, mivel vizuálisan hasonlít a csikóhalra. Ezt a szervet kezdetben inkább a szaglással hozták összefüggésbe, mint a memória elsajátításában betöltött ismert funkciójával. Az orosz Vlagyimir Bekhterev 1900 körül figyelt fel a hippokampusz emlékezetben betöltött szerepére, egy mély memóriazavarban szenvedő beteg megfigyelései alapján. Sokáig azonban a hippokampuszról az volt a hagyományos nézet, hogy a limbikus rendszer többi részéhez hasonlóan az érzelmekért felelős.
A hippokampusz emlékezetben betöltött szerepére a HM beteg hívta fel a kutatók figyelmét. HM számos anterográd és időbeli fokozatú retrográd memóriakárosodásban szenvedett (ilyen károsodásokról szól a Memento című film), miután a gyakori epilepsziás rohamok enyhítése érdekében kétoldalt eltávolították a különböző medialis-temporális lebeny struktúrákat (beleértve a hippokampusz kétoldali ablációját). Különösen fontos, hogy HM továbbra is képes volt eljárási feladatok tanulására (amelyek a striatumhoz kapcsolódnak), és átlagon felüli IQ-val rendelkezett. HM az intelligencia és a deklaratív memória között szembetűnő egyoldali disszociációt mutatott. A hippokampusz-képződés relatív mérete az agy teljes térfogatához viszonyítva gyakran konzerválódik a legtöbb emlősfajban. Mindazonáltal azt találták, hogy ezek a területek a cetfélékben viszonylag hipotrófiásak.
Anatómia
Fő cikk: Hippocampus anatómiája
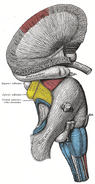
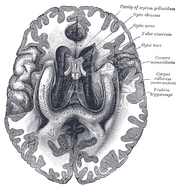
Emberi hippocampus.
Nissl-festett koronális metszet egy makákó majom agyából, a hippocampus (bekarikázva) látható. Forrás: brainmaps.org
Anatómiailag a hippokampusz a cortex szélének kidolgozása. Olyan zónaként különböztethető meg, ahol az agykéreg egyetlen, nagyon sűrűn elhelyezkedő neuronréteggé szűkül, amely szűk S alakúvá görbül. Az agykéreg szélét szegélyező struktúrák alkotják az úgynevezett limbikus rendszert (latin limbus = határ): ezek közé tartozik a hippokampusz, a cinguláris kéreg, a szaglókéreg és az amygdala. Paul MacLean egyszer a hármas agy elméletének részeként felvetette, hogy a limbikus struktúrák tartalmazzák az érzelmek idegi alapjait. A legtöbb idegtudós azonban már nem hisz abban, hogy az egységes “limbikus rendszer” koncepciója érvényes.
A hippokampusz, mint egész, végül úgy néz ki, mint egy ívelt cső, amelyet különbözőképpen analógizáltak egy csikóhalra, vagy egy kos szarvára (Cornu Ammonis), vagy egy banánra.Ez az általános elrendezés az emlősfajok teljes skáláján érvényes, a süntől az emberig, bár a részletek eltérőek. A patkányoknál a két hippokampusz meglepően úgy néz ki, mint egy pár banán, amelyek a száruknál csatlakoznak egymáshoz. Az emberi vagy majomagyban a hippokampusz alsó, a halántéklebeny alapjához közeli része sokkal szélesebb, mint a felső rész. Ennek az összetett geometriának az egyik következménye, hogy a hippokampuszon átvezető keresztmetszetek a vágás szögétől és helyétől függően zavarba ejtően sokféle alakot mutathatnak.
A hippokampusz legerősebb kapcsolatai a mellette a halántéklebenyben elhelyezkedő entorhinális kéreggel (EC) vannak. Az EC felszíni rétegei adják a legtöbb bemenetet a hippokampusznak, és az EC mély rétegei kapják a legtöbb kimenetet. Az EK viszont szoros és kölcsönös kapcsolatban áll az agykéreg számos más részével. A hippokampusz a medialis szeptális területről is nagyon fontos projekciót kap. A szeptális terület elpusztítása megszünteti a hippokampusz théta ritmusát, és súlyosan károsítja a memória bizonyos típusait. (Az úgynevezett “randidrogok” feltehetően legalább részben a medialis septumból a hippokampuszba irányuló kolinerg projekció antagonizálásával fejtik ki amnesztikus hatásukat.)
Fiziológia
Példák patkányok hippokampusz EEG-jéről és CA1 neurális aktivitásáról théta (éber/viselkedő) és LIA (lassú hullámú alvás) módban. Mindegyik ábrán 20 másodpercnyi adat látható, felül egy hippokampusz EEG nyomvonal, középen 40 egyidejűleg rögzített CA1 piramissejtből származó tüske raszterek (minden rasztervonal egy másik sejtet képvisel), alul pedig a futási sebesség ábrája. A felső grafikon egy olyan időszakot ábrázol, amikor a patkány aktívan kereste a szétszórt táplálékgolyókat. Az alsó ábrán a patkány aludt.
A hippokampusz két fő “aktivitási módot” mutat, amelyek mindegyike az EEG-hullámok és a neurális populáció aktivitásának különálló mintázatához kapcsolódik.Ezeket a módokat a hozzájuk kapcsolódó EEG-mintákról nevezték el: théta és nagy szabálytalan aktivitás (LIA). Íme néhány fő jellemzőjük a patkányban, az állatban, amelyet a legkiterjedtebben tanulmányoztak:
A théta-mód az aktív, éber viselkedés (különösen a mozgás) és a REM (álmodó) alvás során is megjelenik. A théta üzemmódban az EEG-t nagy, szabályos hullámok uralják, 6-9 Hz frekvenciatartományban, és a hippokampusz neuronok fő csoportjai (piramissejtek és szemcsesejtek) ritka populációs aktivitást mutatnak, ami azt jelenti, hogy bármely rövid időintervallumban a sejtek nagy többsége néma, míg a fennmaradó kis töredék viszonylag nagy sebességgel tüzel, a legaktívabbaknál akár 50 tüske is lehet egy másodperc alatt. Egy aktív sejt jellemzően fél másodperctől néhány másodpercig marad aktív. Ahogy a patkány viselkedik, az aktív sejtek elnémulnak, és új sejtek válnak aktívvá, de az aktív sejtek teljes százalékos aránya többé-kevésbé állandó marad. Sok helyzetben a sejtaktivitást nagymértékben meghatározza az állat térbeli elhelyezkedése,de más viselkedési változók is egyértelműen befolyásolják.
A LIA-mód lassú hullámú (nem álmodó) alvás során, valamint az éber mozdulatlanság állapotaiban, például pihenés vagy evés közben is megjelenik. A LIA módban az EEG-t az éles hullámok uralják, amelyek az EEG jel 200-300msec-ig tartó, véletlenszerűen időzített nagy eltérései. Ezek az éles hullámok határozzák meg a populációs idegi aktivitásmintákat is. Közöttük a piramissejtek és a szemcsesejtek nagyoncsendesek (de nem némák). Egy éles hullám alatt a populáció 5-10%-a 50 msec alatt akciós potenciálokat bocsát ki; e sejtek közül sokan nem egy, hanem tüskék sorozatát bocsátják ki.
Ez a két hippokampusz aktivitási mód a főemlősökben és a patkányokban is megfigyelhető, azzal a fontos kivétellel, hogy a főemlős hippokampuszban nehéz volt robusztus théta ritmizmust látni. Vannak azonban minőségileg hasonló éles hullámok, és hasonlóállapotfüggő változások az idegi populációaktivitásban..
A théta ritmus
A sűrűn elhelyezkedő idegi rétegek miatt a hippokampusz generálja az egyik legnagyobb EEG jeleket bármely agyi struktúra közül. Bizonyos helyzetekben az EEG-t szabályos hullámok uralják, amelyek gyakran több másodpercig tartanak. Ezt az EEG-mintát théta ritmusnak nevezik. Ez volt az egyik legkorábban felfedezett EEG-jelenség: az első leírás Jungtól és Kornmullertől származik, 1938-ban. Az érdeklődés azonban csak 1954-ben, Green és Arduini nyulakon, macskákon és majmokon végzett hosszú és alapos tanulmányának publikálásával indult meg igazán.Talán nagyrészt azért, mert a théta ritmust az akkoriban aktuális témának számító izgalommal hozták összefüggésbe, tanulmányuk a következő tanulmányok áradatát váltotta ki, aminek eredményeként az 1950-es és 1960-as években szó szerint több száz tanulmány jelent meg a théta ritmus fiziológiájáról és farmakológiájáról. E meglehetősen ijesztő mennyiségű munka ellenére sok kérdés megválaszolatlan maradt, különösen a funkció kérdése. Még jelenleg sem sikerült meggyőzően megválaszolni ezt a legkritikusabb kérdést.
A théta ritmikusság nagyon nyilvánvaló a nyulakban és a rágcsálókban, és egyértelműen jelen van a macskákban és a kutyákban is. Az, hogy a théta látható-e az ősállatokban, kínzó kérdés. Green és Arduini csak nagyon rövid, meglehetősen szabálytalan ritmikus kitörésekről számolt be majmoknál, és a legtöbb későbbi vizsgálat ennél többet nemigen látott. A módszertani eltérések azonban megnehezítették az erős következtetések levonását.
Patkányoknál (a messze a legtöbbet vizsgált állatoknál) a théta főként két állapotban figyelhető meg: először, amikor az állat sétál vagy más módon aktívan interakcióba lép a környezetével; másodszor, a REM-alvás során. A frekvencia a futási sebesség függvényében nő, az alacsony tartományban körülbelül 6,5 Hz-nél kezdődik, és a magas tartományban körülbelül 9 Hz-ig emelkedik, bár néha magasabb frekvenciák is megfigyelhetők drámai mozgások, például nagy távolságokon való átugrás esetén. Más, nagyobb állatfajoknál a thetafrekvenciák általában valamivel alacsonyabbak. Úgy tűnik, hogy a viselkedési függőség is fajonként változik: macskáknál és nyulaknál a thétafrekvencia gyakran megfigyelhető a mozdulatlan éberség állapotában. Ezt patkányok esetében is megfigyelték, de csak akkor, ha súlyosan megijedtek.
A théta nem csak a hippokampuszra korlátozódik. Patkányoknál az agy számos részén megfigyelhető, beleértve szinte mindazokat, amelyek erősen kölcsönhatásban vannak a hippokampusszal. Úgy gondolják, hogy a ritmus pacemakere a medialis szeptális területen található: ez a terület az összes olyan régióra vetül, amely théta ritmikusságot mutat, és pusztulása megszünteti a thetát az egész agyban. (Talán van egy kivétel, egy kis terület a hipotalamuszban, az úgynevezett supramamillarynucleus, amely úgy tűnik, hogy bizonyos helyzetekben képes fenntartani a theta-t a septumtól függetlenül.)
A theta funkcióját, ha feltételezzük, hogy van funkciója, még nem sikerült meggyőzően megmagyarázni, bár számos elméletet javasoltak már.A legnépszerűbb irányzat a tanulással és a memóriával kapcsolja össze.Jól ismert, hogy a középső szeptum – a théta rendszer központi csomópontjának – sérülése súlyos memóriazavarokat okoz.A középső szeptum azonban több mint a théta irányítója,ahippocampusba irányuló kolinerg projekciók fő forrása is. Nem igazolták, hogy a szeptális sérülések kifejezetten a theta megszüntetésével fejtenék ki hatásukat.
Éles hullámok
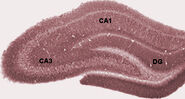
A hippokampusz EEG-je alvás közben vagy ébrenléti állapotokban, amikor az állat pihen vagy más módon nem foglalkozik a környezetével, szabálytalan lassú hullámok mintázatát mutatja, amelyek amplitúdója valamivel nagyobb, mint a theta-hullámoké. Ezt a mintázatot időnként nagy hullámok, úgynevezett éles hullámok szakítják meg. Ezek az események a CA3 ésCA1 piramissejtjeiben 50-100 msec időtartamú tüskeaktivitás-robbanásokhoz kapcsolódnak. Rövid ideig tartó, nagyfrekvenciás EEG-ingadozásokhoz is társulnak, amelyeket “hullámzásnak” neveznek. A patkányoknál 150-200 Hz közötti frekvenciájú hullámzásokat általában csak a CA1 sejttestrétegben vagy ahhoz nagyon közel elhelyezkedő elektródákkal lehet észlelni. Ezzel szemben a hippokampuszon belül bárhol, vagy akár a szomszédos agyi struktúrákban elhelyezett elektródák gyakran érzékelik az éles hullámokat, mint nagyméretű, lassú, 200-400 msec időtartamú EEG-eltéréseket.
Patkányoknál az éles hullámok alvás közben a legerősebbek, amikor másodpercenként átlagosan 1 körüli frekvenciával, de nagyon szabálytalan időbeli mintázatban jelentkeznek. Az éles hullámok inaktív ébrenléti állapotokban is előfordulnak, de ilyenkor ritkábbak és általában kisebbek. Éles hullámokat figyeltek meg az emberi halántéklebenyben és a majom hippokampuszában is.Majmokban az éles hullámok meglehetősen erősek, de közel sem fordulnak elő olyan gyakran, mint patkányokban.
Az éles hullámok egyik legérdekesebb aspektusa az, hogy úgy tűnik, hogy kapcsolatban állnak a memóriával. Wilson és McNaught 1994-ben és számos későbbi tanulmányban arról számolt be, hogy amikor a hippokampusz helysejtjei átfedő térbeli tüzelési mezőkkel rendelkeznek (és ezért gyakran közel egyidejűleg tüzelnek), hajlamosak korrelált aktivitást mutatni a viselkedési ülést követő alvás során. A korrelációnak ezt a fokozódását, amelyet általában reaktivációnak neveznek, főként az éles hullámokra korlátozódónak találták. Azt javasolták, hogy az éles hullámok valójában a viselkedés során megjegyzett neurális aktivitási minták reaktivációi, amelyeket a szinaptikus kapcsolatok megerősödése vezérel a hippokampuszon belül. Ez az elképzelés a Buzsaki és mások által képviselt “kétlépcsős memória” elmélet egyik kulcseleme, amely szerint az emlékek a hippokampuszban tárolódnak a viselkedés során, majd később alvás közben átkerülnek a neokortexbe: az éles hullámok feltételezhetően a hippokampusz kimeneti pályáinak neokortikális célpontjaiban bekövetkező Hebb-féle szinaptikus változásokat hajtják.
Evolúció
A hippokampusz általában hasonló megjelenésű az emlősfajok között, a bazális fajoktól, mint a süntől a leg “fejlettebbekig”, mint az ember.A hippokampusz mérete a testmérethez viszonyítva nagyjából növekszik, a főemlősöknél körülbelül kétszer akkora, mint a sünnél. A hippokampusz azonban közel sem növekszik olyan ütemben, mint a neokortex-testméret arány.Így a hippokampusz sokkal nagyobb térfogatot foglal el a rágcsálók agykérgéből, mint a főemlősöknél.
A hippokampusz mérete és a térbeli memória között is van egy általános kapcsolat: amikor hasonló fajok közötti összehasonlítást végeznek, a nagyobb térbeli memóriaképességgel rendelkező fajok általában nagyobb hippokampusz térfogattal rendelkeznek. Ez az összefüggés a nemi különbségekre is kiterjed: azoknál a fajoknál, ahol a hímek és nőstények erős különbségeket mutatnak a térbeli emlékezőképességben, a hippokampusz térfogatában is hajlamosak ennek megfelelő különbségeket mutatni
A nem emlős fajoknak nincs az emlősök hippokampuszára hasonlító agyi struktúrájuk, de van egy olyan, amely homológnak tekinthető vele. A hippokampusz, amint arra fentebb rámutattunk,lényegében az agykéreg mediális szegélye. Csak az emlősöknek van teljesen kifejlett agykéregük, de a szerkezet, amelyből fejlődött ki, az úgynevezett pallium, minden gerincesben jelen van, még a legprimitívebbekben is, mint például a lámpás vagy a süllő. A palliumot általában három zónára osztják: mediális, laterális és dorzális. A mediális pallium alkotja a hippokampusz előfutárát. Vizuálisan nem hasonlít a hippokampuszra, mivel a rétegek nem S alakúvá torzultak, és a fogazott gyrus sem burkolja be őket, de a homológiát erős kémiai és funkcionális rokonság jelzi. Ma már bizonyíték van arra, hogy ezek a hippokampusz-szerű struktúrák részt vesznek a térbeli megismerésben madaraknál, hüllőknél és halaknál.
A madaraknál a megfelelés elég jól megalapozott ahhoz, hogy a legtöbb anatómus a medialis pallialis zónát “madárhippokampusznak” nevezi. Számos madárfaj erős térbeli képességekkel rendelkezik,különösen azok, amelyek élelmet rejtegetnek. Bizonyított, hogy a táplálékot rejtegető madaraknak nagyobb a hippokampuszuk, mint más madárfajoknak, és hogy a hippokampusz károsodása a térbeli memória károsodását okozza.
A halak esetében a történet összetettebb. A teleost halaknál (amelyek a létező fajok nagy többségét alkotják) az előagy furcsán eltorzult más gerincesekhez képest. A legtöbb neuroanatómus úgy véli, hogy a teleoszthal előagya lényegében kifordított, mint egy kifordított zokni, így azok a struktúrák, amelyek a legtöbb gerincesnél belül, a kamrák mellett helyezkednek el, a teleoszthalnál kívül találhatók, és fordítva. Ennek egyik következménye, hogy a tipikus gerincesek mediális palliumáról (“hippokampusz” zónája) azt gondolják, hogy megfelel a tipikus halak laterális palliumának. Számos halfajról (különösen az aranyhalakról) kísérleti úton kimutatták, hogy erős térbeli emlékezőképességgel rendelkeznek, sőt “kognitív térképeket” alkotnak az általuk lakott területekről. Bizonyítékok vannak arra, hogy az oldalsó pallium károsodása károsítja a térbeli emlékezetet (a hosszú távú navigáció, mint például a lazacok tájékozódása, azonban úgy tűnik, más mechanizmusokra támaszkodik.)
A hippokampusz régió navigációban betöltött szerepe tehát úgy tűnik, hogy a gerincesek evolúciójában jóval korábban kezdődött, megelőzve a több százmillió évvel ezelőtt bekövetkezett szétválást. Még nem ismert, hogy a medialis pallium hasonló szerepet játszik-e a még kezdetlegesebb gerinceseknél, például a cápáknál és a rájáknál, vagy akár a lámpáshalaknál és a süllőhalaknál. Egyes rovarok és puhatestűek, mint például a polipok, szintén erős térbeli tanulási és navigációs képességekkel rendelkeznek, de úgy tűnik, hogy ezek az emlősök térbeli rendszerétől eltérően működnek, így egyelőre nincs jó okunk azt gondolni, hogy közös evolúciós eredetük van; és az agyszerkezetben sincs elegendő hasonlóság ahhoz, hogy ezekben a fajokban bármi, ami a “hippokampuszra” hasonlítana, azonosítható legyen.
Lásd még
- Amygdalohippocampectomia
- Medián elülső agyköteg
- Septum. magok
Hivatkozások & Bibliográfia
Főbb szövegek
Könyvek
- Aboitiz, F, Morales D, Montiel J (2003). Az emlősök izokortexének evolúciós eredete: Egy integrált fejlődési és funkcionális megközelítés felé. Behav. Brain Sciences 26: 535-552.
- Amaral, D; Lavenex P (2006). “Ch 3. Hippocampal Neuroanatomy” Andersen P, Morris R, Amaral D, Bliss T, O’Keefe J The Hippocampus Book, Oxford University Press.
- Broglio, C, Gómez A, Durán E, Ocaña FM, Jiménez-Moya F, Rodríguez F, Salas C (2002). Egy közös előagyi gerinces terv jellegzetességei: Specializált palliális területek a térbeli, időbeli és érzelmi emlékezethez az aktinopterygiai halakban. Brain Res. Bull. 57: 397-399.
- Buzsáki, G (2002). Théta oszcillációk a hippokampuszban. Neuron 33: 325-340.
- Buzsáki, G (2006). Rhythms of the Brain.
- Cantero, JL, Atienza M, Stickgold R, Kahana MJ, Madsen JR, Kocsis B (2003). Alvásfüggő théta oszcillációk az emberi hippokampuszban és neokortexben. J. Neurosci. 23: 10897-10903.
- Colombo, M, Broadbent N (2000). A madárhippokampusz az emlős hippokampusz funkcionális homológja? Neurosci. Biobehav. Rev. 24: 465-484.
- Eichnbaum, H; Cohen NJ (1993). Memory, Amnesia, and the Hippocampal System, MIT Press.
- Ekstrom, AD, Kahana MJ, Caplan JB, Fields TA, Isham EA, Newman EL, Fried I (2003). Az emberi térbeli navigáció alapjául szolgáló sejthálózatok. Nature 425: 184-188.
- Gray, JA; McNaughton N (2000). A szorongás neuropszichológiája: An Enquiry into the Functions of the Septo-Hippocampal System, Oxford University Press.
- Green, JD, Arduini AA (1954). A hippokampusz elektromos aktivitása arousalban. J. Neurophysiol. 17: 533-557.
- Jacobs, LF, Gaulin SJ, Sherry DF, Hoffman GE (1990). A térbeli megismerés evolúciója: a térbeli viselkedés nemspecifikus mintái előre jelzik a hippokampusz méretét. PNAS 87: 6349-6352.
- Jacobs, LF (2003). A kognitív térkép evolúciója. Brain Behav. Evol. 62: 128-139.
- Kirk, IJ, McNaughton N (1991). A szupramammilláris sejtek tüzelése és a hippokampusz ritmikus lassú aktivitása. Neuroreport 11: 723-725.
- Maguire, EA, Gadian DG, Johnsrude IS, Good CD, Ashburner J, Frackowiak RS, Frith CD (2000). Navigációval kapcsolatos szerkezeti változások a taxisok hippocampijában. PNAS 97: 4398-4403.
- McNaughton, BL, Battaglia FP, Jensen O, Moser EI, Moser MB (2006). Az útvonal-integráció és a “kognitív térkép” neurális alapja. Nat. Rev. Neurosci. 7: 663-678.
- Moser, EI, Kropf E, Moser M-B (2008). Helysejtek, rácssejtek és az agy térbeli reprezentációs rendszere. Ann. Rev. Neurosci. 31.
- Nieuwenhuys, R (1982). An Overview of the Organization of the Brain of Actinopterygian Fishes. Am. Zool. 22: 287-310.
- O’Kane, G, Kensinger EA, Corkin S (2004). Bizonyíték a szemantikai tanulásra mély amnéziában: Egy vizsgálat H.M. beteggel. Hippocampus 14: 417-425.
- O’Keefe, J; Nadel L (1978). The Hippocampus as a Cognitive Map, Oxford University Press.
- Portavella, M, Vargas JP, Torres B, Salas C (2002). A telencephalicus pallialis sérülések hatása a térbeli, időbeli és érzelmi tanulásra aranyhalakban. Brain Res. Bull. 57: 397-399.
- Rodríguez, F, Lópeza JC, Vargasa JP, Broglioa C, Gómeza Y, Salas C (2002). A térbeli emlékezet és a hippokampusz pallium a gerincesek evolúciója során: meglátások a hüllők és a teleosztrák halaiból. Brain Res. Bull. 57: 499-503.
- Sainsbury, RS, Heynen A, Montoya CP (1987). A hippokampusz 2. típusú théta viselkedési korrelátumai patkányban. Physiol. Behav. 39: 513-519.
- Scoville, WB, Milner B (1957). A közelmúltbeli emlékezet elvesztése kétoldali hippokampusz sérülések után. J. Neurol. Neurosurg. Psych. 20: 11-21.
- Shettleworth, SJ (2003). Memória és hippokampusz specializáció a táplálékot tároló madaraknál: Kihívások az összehasonlító kogníció kutatásához. Brain Behav. Evol. 62: 108-116.
- Skaggs, WE, McNaughton BL, Wilson MA, Barnes CA (1996). Théta fázis precesszió a hippokampusz neuronpopulációkban és az időbeli szekvenciák tömörítése. Hippocampus 6: 149-176.
- Squire, LR (1992). A memória és a hippokampusz: a patkányokkal, majmokkal és emberekkel végzett vizsgálatok eredményeinek szintézise. Psych. Rev. 99: 195-231.
- Squire, LR; Schacter DL (2002). The Neuropsychology of Memory, Guilford Press.
- Vanderwolf, CH (1969). A hippokampusz elektromos aktivitása és az önkéntes mozgás patkányban. EEG & Clin. Neurophysiol. 26: 407-418.
- Vargas, JP, Bingman VP, Portavella M, López JC (2006). Telencephalon és geometriai tér az aranyhalakban. Eur. J. Neurosci. 24: 2870-2878.
- West, MJ (1990). A hippokampusz sztereológiai vizsgálatai: különböző fajok, köztük sünök, laboratóriumi rágcsálók, vad egerek és emberek hippokampusz alosztályainak összehasonlítása. Prog. Brain Res. 83: 13-36.
Papers
- Amaral DG és Cowan WM. 1980. A hippocampus képződmény szubkortikális afferensei a majomban. Journal of Comparative Neurology. Feb 15; 189(4):573-91.
- Duvernoy, H. (2005) The Human Hippocampus, 3rd ed. Berlin: Springer-Verlag.
További anyagok
Lapok
- Google Scholar
- http://www.psycheducation.org/emotion/hippocampus.htm Nagyszerű agytúrák!!!
- BrainMaps at UCDavis hippocampus
- Artificial Hippocampus
- BrainInfo at the University of Washington hier-164
- John O’Keefe & Lynn Nadel (1978) The Hippocampus as a Cognitive Map , Oxford University Press. Teljes szöveg
További képek
Fotó hozzáadása a galériához
Az emberi agy: Limbikus rendszer
Amygdala – Cinguláris gyrus – Fornicus gyrus – Hippocampus – Hypothalamus – Mammilláris test – Nucleus accumbens – Orbitofrontális kéreg – Parahippocampus gyrus
|
v-d-e
Emberi agy: (TA A14.1.09.002-240, 301-320, GA 9.818-826) |
|||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Frontális lebeny |
|
||||||||||||||||
| Parietális lebeny |
|
Superolaterális
|
Medial/inferior
|
Mindkettő
|
|||||||||||||
| Occipitális lebeny |
|
Superolaterális
|
Medial/inferior
|
||||||||||||||
| Temporális lebeny |
|
Superolaterális
|
Középső/alsó
|
||||||||||||||
| Interlobaris sulci/fissures |
|
Superolaterális
|
Medial/inferior
|
||||||||||||||
| Limbikus lebeny |
|
Más
|
|||||||||||||||
| Insularis lebeny |
Az insula hosszú gyrusza – Az insula rövid gyrusza – Az insula körkörös sulcusza |
||||||||||||||||
| Általános |
Operculum – Az agyféltekék pólusai |
||||||||||||||||
Egyes kategorizálások közelítések, és néhány Brodmann-terület átfogja a gyri-ket.
|
|||||||||||||||||
Az oldal a Wikipédia Creative Commons licencelt tartalmát használja (szerzők megtekintése).