評価|生物心理学|比較|認知|発達|言語|個人差|性格|哲学|社会|
方法|統計|臨床|教育|産業|専門項目|世界心理|
Biological.Lab:行動遺伝学|進化心理学|神経解剖学|神経化学|神経内分泌学|神経科学|精神神経免疫学|生理心理学|精神薬理学(索引、概要)
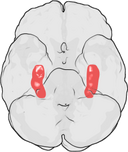
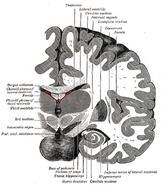
人間の脳における海馬の位置について。
同名の雑誌については、海馬(雑誌)
海馬は前脳の一部で、側頭葉の内側に位置する。 大脳辺縁系に属し、長期記憶と空間ナビゲーションに大きな役割を果たす。 ヒトをはじめとする哺乳類には海馬が2つあり、左右に1つずつあります。 海馬は、最も広く研究されているげっ歯類では、バナナのような形をしている。 ヒトの場合、海馬は湾曲した複雑な形をしており、初期の解剖学者たちはタツノオトシゴを思い浮かべたという。
アルツハイマー病では、海馬は脳の中で最初に損傷を受ける領域の1つで、記憶障害や方向感覚の喪失が最初の症状として現れます。 海馬の損傷は、酸素欠乏(無酸素状態)、脳炎、中側頭葉てんかんでも起こります。
海馬の機能
おそらく最も古い考えは、海馬が嗅覚に関与しているというもので、これは主に脳内で嗅覚皮質の隣に位置していることから示唆されたようである。
長年にわたり、海馬の機能については、抑制、記憶、空間という3つの主要な考え方が文献を支配してきた。 行動抑制説(オキーフとナデルは「ブレーキを踏め!」と戯画化した)は、1960年代まで非常に人気があった。 第一に、海馬に損傷を受けた動物は多動になる傾向があること、第二に、海馬に損傷を受けた動物は以前に教えられた反応を抑制することを学ぶのが困難であることが多いことであった。 ジェフリー・グレイは、この考えを発展させ、不安における海馬の役割に関する本格的な理論へと発展させた。 しかし、この抑制理論は現在ではあまり人気がない。
2番目の重要な考え方は、海馬と記憶との関連である。 その結果、H.M.は手術後やその数年前に起こった出来事を意識的に思い出すことができなくなったのである。 H.M.は、史上最も多く研究された医学的症例といわれるほど、この症例は大きな関心を呼んだ。 その後、海馬の損傷や記憶喪失(事故や病気による)が同程度の他の患者も研究され、文字通り何千もの実験によって海馬の神経可塑性の生理学が研究されてきた。 現在では、海馬が記憶において何らかの重要な役割を担っていることはほぼ共通している。しかし、この役割の正確な性質については、依然として広く議論されている。
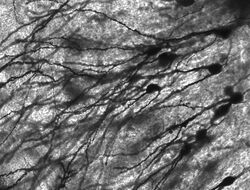
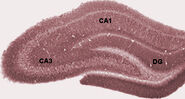
ネズミの海馬のゴルジ染色されたニューロン
Role in memory
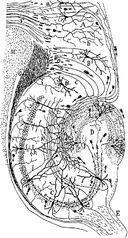
ネズミの海馬の神経回路の図面。 S. Ramón y Cajal, 1911.
心理学者や神経科学者は、海馬の正確な役割について論争しているが、一般的には、海馬が個人的に経験した出来事に関する新しい記憶(エピソード記憶または自伝的記憶)の形成に不可欠な役割を持つことに同意している。 海馬は、より大きな内側側頭葉の記憶システムの一部として、一般的な宣言的記憶(明示的に言語化できる記憶-例えば、エピソード記憶に加えて事実に関する記憶も含まれる-)に関与すると考える研究者もいる。 記憶における海馬の役割
空間記憶とナビゲーションにおける役割
海馬が空間情報の保存と処理に関与していることを示す証拠がいくつかある。 ラットの研究により、海馬の神経細胞は空間発火野を持つことが示されている。 これらの細胞は場所細胞と呼ばれる。 一部の細胞は、動物が特定の場所にいると、進行方向に関係なく発火するが、大部分は頭の方向と進行方向に少なくとも部分的に敏感である
。 空間記憶とナビゲーションにおける海馬の役割
海馬とうつ病
- 海馬の体積減少、おそらく過剰なグルココルチコイドの
神経発生への影響を反映していることが、対照群と比較してうつ病患者群で確認されている
主要記事。 うつ病と海馬
歴史
解剖学者Giulio Cesare Aranzi(1564年頃)が、タツノオトシゴに視覚的に似ていることから、海馬という言葉を初めて大脳の器官に使ったとされる。 この器官は当初、記憶の獲得という機能よりも、むしろ嗅覚と結びつけられていた。 1900年頃、ロシアのウラジーミル・ベクテレフが、重度の記憶障害を持つ患者を観察し、記憶における海馬の役割に言及した。 しかし、長年にわたり、海馬は大脳辺縁系の他の部分と同様に情動を司るというのが従来の見解だった。
記憶における海馬の重要性は、患者HMによって研究者の注意を引くことになった。 HMは、頻発するてんかん発作を緩和するために、様々な内側側頭葉の構造を両側から切除(両側の海馬の切除を含む)した後、多くの前向きの記憶障害と時間的に勾配した逆向きの記憶障害(こうした障害は映画『メメント』の主題となっている)に悩まされた。 特に重要なのは、HMが依然として(線条体と関連する)手続き的課題を学習することができ、平均以上のIQを持っていたことである。 HMは、知能と宣言的記憶の間に顕著な単一解離があることを実証した。 海馬の相対的な大きさは、哺乳類のほとんどの種で、脳の総体積との関係で保存されていることが多い。 それにもかかわらず、鯨類ではこれらの領域が比較的低栄養であることが分かっている
解剖学
主な記事 海馬の解剖学
ヒトの海馬.
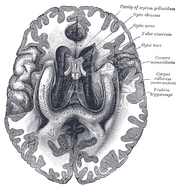
マカクザルの脳のニッスル染色冠状断、丸付きは海馬の様子である。 Source: brainmaps.org
解剖学的に、海馬は大脳皮質の端の精巧な部分である。 海馬は、皮質が狭くなり、神経細胞が非常に密に詰まった1つの層となり、S字型に丸まったゾーンとして区別できます。 大脳皮質の縁に並ぶ構造は、いわゆる辺縁系(ラテン語でlimbbus =境界)を構成する。これらには海馬、帯状皮質、嗅覚皮質、および扁桃体が含まれる。 ポール・マクリーンはかつて、三位一体脳説の一環として、大脳辺縁系が情動の神経基盤を構成していることを示唆した。
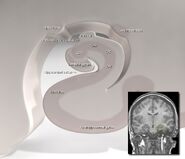
海馬は全体として湾曲したチューブのような形をしており、タツノオトシゴ、雄羊の角(Cornu Ammonis)、バナナなどさまざまに例えられる。このレイアウトはハリネズミからヒトまで、哺乳類のすべての種で共通しているが、細部は異なっている。 ラットの場合、2つの海馬は茎でつながった一対のバナナのように見えるのが不思議だ。 ヒトやサルの脳では、海馬の下部、側頭葉の基部に近い部分は、上部にある部分よりずっと広い。 この複雑な形状の結果、海馬の断面は、切る角度や位置によって、実にさまざまな形状を示す。
海馬は、側頭葉で隣接する内耳皮質(EC)と最も強く結合している。 ECの表層は海馬への最も多くの入力を提供し、ECの深層は最も多くの出力を受け取る。 ECは大脳皮質の他の多くの部分と相互に強く結合している。 海馬はまた、内側中隔領域から非常に重要な投射を受けている。 この中隔領域を破壊すると、海馬のシータリズムが消失し、ある種の記憶が著しく損なわれる。 (いわゆる「デートレイプ」薬は、少なくとも部分的には内側中隔から海馬へのコリン作動性投射に拮抗することによって、記憶阻害作用を示すと考えられている。)
生理学
ラット海馬の脳波とCA1神経活動のシータ(覚醒/行動)モードとLIA(徐波睡眠)モードの例です。 各プロットは20秒間のデータを示し、上部は海馬EEGトレース、中央は同時に記録した40個のCA1錐体細胞からのスパイクラスタ(ラスタ線はそれぞれ異なる細胞を表す)、下部は走行速度のプロットである。 上段のプロットは、ラットが散乱した餌を活発に探していた時間帯を表している。
海馬は2つの主要な活動モードを示し、それぞれが脳波と神経集団の活動の明確なパターンと関連している。これらのモードは、それらに関連する脳波パターンから名付けられた:シータと大不規則活動(LIA)。 シータモードとは、活動的で警戒心の強い行動(特に運動)をとっている時や、レム睡眠(夢を見ている時)に出現するモードである。 シータモードでは、脳波は6~9Hzの大きな規則的な波で支配され、海馬の主要なニューロン群(錐体細胞および顆粒細胞)は疎な集団活動を示す。 活動中の細胞は通常半秒から数秒間活動したままである。 ラットが行動するにつれ、活性細胞は沈黙し、新しい細胞が活性化するが、全体の活性細胞の割合はほぼ一定に保たれる。 多くの場合、細胞活性は動物の空間的位置によって大きく左右されるが、他の行動変数も明らかに影響を与える。
LIAモードは徐波(非夢想)睡眠中や、休憩や食事など覚醒時の無動状態にも現れる。 LIAモードでは、脳波は鋭波に支配される。鋭波は、200~300ミリ秒の脳波信号のランダムなタイミングでの大きな偏向を示すものである。 この鋭角波が母集団の神経活動パターンを決定している。 この間、錐体細胞や顆粒細胞は非常に静かである(無音ではない)。
これら2つの海馬の活動様式はラットだけでなく霊長類にも見られるが、霊長類の海馬では強固なシータリズムを見ることが困難であるという重要な例外を除いては、シータリズムを見ることができる。
シータリズム
海馬は神経層が密集しているため、あらゆる脳構造の中で最も大きな脳波信号を発生させることができる。 ある状況下では、脳波は規則的な波で支配され、しばしば何秒にもわたって継続する。 この脳波のパターンはシータリズムとして知られている。 シータリズムは、1938年にJungとKornmullerによって発見された最も初期の脳波現象の一つである。 しかし、1954年にグリーンとアルドゥイーニがウサギ、ネコ、サルにおけるシータリズムの長期にわたる徹底的な研究を発表して以来、その関心は急速に高まった。彼らの論文は、当時話題となっていたシータリズムを覚醒と関連づけたことが大きな要因となって、後続研究が殺到し、1950年代と1960年代にシータの生理学と薬理学の研究が文字通り数百件発表されるに至った。 このように、かなり膨大な量の研究が行われたにもかかわらず、多くの疑問、特に機能に関する疑問は未解決のままであった。 655>
シータリズムはウサギやげっ歯類では非常に明白であり、ネコやイヌでも明らかに存在する。 霊長類にシータが見られるかどうかは、悩ましい問題である。 GreenとArduiniはサルにおいて非常に短いバースト状の不規則なリズムを報告しただけであり、その後のほとんどの研究ではそれ以上のものは見られていない。
ラット(最も広範囲に研究されている動物)では、シータは主に次の2つの条件で見られる:第1に、動物が歩いているとき、または他の方法で周囲と活発に相互作用しているとき、第2に、レム睡眠中である。 その周波数は走行速度の関数として増加し、低い方で約6.5Hz、高い方で約9Hzまで増加するが、広い隙間を飛び越えるような劇的な動きの場合にはより高い周波数が見られることもある。 他の大型の動物種では、シータ周波数は一般にもう少し低い。 行動依存性も動物種によって異なるようで、ネコやウサギでは、動きのない覚醒状態のときにシータがよく観察される。 これはラットでも同様に報告されているが、ひどく怯えているときのみである
シータは海馬だけに限定されるものではない。 ラットでは、海馬と強く相互作用するほぼすべての部位を含む、脳の多くの部位で観察することができる。 このリズムのペースメーカーは内側中隔領域にあると考えられており、この領域はシータリズムを示すすべての領域に投射しており、この領域を破壊すると脳全体のシータが消失する。 (
シータの機能は、それが1つを持っていると仮定すると、多くの理論が提案されているが、まだconvincently説明されていません。しかし、中隔は単にシータを制御するだけでなく、海馬へのコリン作動性投射の主要な供給源でもある。
鋭い波
睡眠中、あるいは動物が休息しているか、さもなければ周囲の環境に関与していない覚醒状態では、海馬のEEGは、θ波よりやや大きな振幅の不規則な徐波のパターンを示す。 このパターンは時折、鋭敏波と呼ばれる大きな波で中断される。 これらの事象は、CA3およびCA1の錐体細胞における50-100ミリ秒持続するスパイク活動のバーストと関連している。 また、「波紋」と呼ばれる短時間の高周波の脳波振動を伴うこともある。 ラットでは150-200Hzの周波数を持つ波紋は、通常、CA1細胞体層の内部またはそのごく近傍にある電極によってのみ検出されうる。 これとは対照的に、海馬の内部や隣接する脳構造のどこかにある電極では、200~400ミリ秒の大きなスロー脳波偏向として鋭い波が検出されることが多い。 鋭敏波は非活動的な覚醒状態でも発生するが、そのときの頻度は低く、通常はより小さい。 サルの場合、鋭角波はかなり強固であるが、ラットほど頻繁には発生しない。
鋭角波の最も興味深い点の1つは、それが記憶と関連しているように思われることである。 海馬の場所細胞が空間的な発火野を重複して持つ場合(したがって、しばしばほぼ同時に発火する)、行動セッションに続く睡眠中に相関した活動を示す傾向があると、WilsonとMcNaughton 1994、およびその後の多くの研究が報告している。 この相関の増強は一般に再活性化と呼ばれ、主に鋭い波に限定されていることが分かっている。 これは、海馬内のシナプス結合が強化され、行動時に記憶された神経活動パターンが再活性化されたものであると考えられています。 この考えは、ブザキらが提唱する「二段階記憶説」の重要な構成要素である。この説では、記憶は行動中に海馬に保存され、その後睡眠中に大脳新皮質に転送されるとする。つまり、シャープ波は海馬出力経路の皮質標的におけるヘブのシナプス変化を引き起こすと考えられる。
Evolution
海馬は、ハリネズミのような底辺の種からヒトのような最も「進んだ」種まで、哺乳類の範囲内で概ね同様の外観を持つ。海馬のサイズ対体格比は広く増加し、霊長類ではハリネズミの約2倍となる。 このように、海馬は霊長類よりも齧歯類の方が皮質の大部分を占めている。
海馬の大きさと空間記憶の間には一般的な関係もあり、同種間で比較した場合、空間記憶の能力が高いものは海馬の体積が大きい傾向がある。 この関係は性差にも及び、雌雄で空間記憶能力に強い差がある種では、海馬の体積にも対応する差が見られる傾向がある
哺乳類以外の種には、哺乳類の海馬に似た脳構造はないが、相同と思われるものがある。 海馬は、上で指摘したように、本質的には大脳皮質の内縁である。 大脳皮質が完全に発達しているのは哺乳類だけだが、そこから進化した「壁」と呼ばれる構造は、すべての脊椎動物、それもヤツメウナギやタガメのような最も原始的な動物にさえ存在する。 通常、口蓋垂は内側、外側、背側の3つのゾーンに分けられる。 内側は海馬の前駆体であり、外側は海馬の前駆体である。 海馬とは、層がS字に歪んでおらず、歯状回に包まれていないため、視覚的には似ていないが、化学的、機能的に強い親和性を示す。 鳥類では、海馬に似た構造が鳥類、爬虫類、魚類の空間認知に関与しているという証拠がある
鳥類では、この対応は十分に確立されており、ほとんどの解剖学者は内側口蓋帯を「鳥類の海馬」と呼んでいる。 鳥類の多くの種は強い空間的能力を持っており、特に餌をキャッシュする種が多い。 また、海馬の損傷は空間記憶の障害を引き起こすという証拠もある。
魚類の話はもっと複雑です。 現存する種の大半を占める魚類のうち望遠魚では、前脳が他の脊椎動物に比べて奇妙なほど歪んでいるのです。 ほとんどの神経解剖学者たちは、前脳の形が靴下を裏返したように逆になっていて、ほとんどの脊椎動物では脳室の隣の内側にある構造が、魚類では外側にあり、その逆もまたしかりだと考えています。 その結果、脊椎動物の内側葉状体(海馬部)は、魚類の外側葉状体に相当すると考えられている。 魚類(特に金魚)には強い空間記憶能力があることが実験的に示されており、生息する地域の「認知地図」を形成することさえある。 しかし、サケのホーミングのような長距離ナビゲーションは、別のメカニズムに依存しているようだ)
したがって、ナビゲーションにおける海馬領域の役割は、脊椎動物の進化のはるか昔、数億年前に起こった分裂より前に始まったようである。 サメやエイ、あるいはウナギやタガメなど、さらに原始的な脊椎動物において、内側頭蓋が同様の役割を果たしているかどうかはまだ分かっていない。 昆虫やタコなどの軟体動物にも強力な空間学習能力やナビゲーション能力があるが、これらは哺乳類の空間システムとは異なる働きをするようで、進化の過程で共通の起源を持つと考える十分な根拠はまだない。
- Amygdalohippocampectomy
- Median forebrain bundle
- Septal(海馬)の項も参照してください。 nuclei
References & Bibliography
Key texts
Books
- Aboitiz, F, Morales D, Montiel J (2003). 哺乳類の等角皮質の進化的起源。 発達と機能の統合的なアプローチに向けて。 Behav. 脳科学26:535-552.
- Amaral, D; Lavenex P (2006). “Ch 3. Hippocampal Neuroanatomy” Andersen P, Morris R, Amaral D, Bliss T, O’Keefe J The Hippocampus Book, Oxford University Press.
- Broglio, C, Gómez A, Durán E, Ocaña FM, Jiménez-Moya F, Rodríguez F, Salas C (2002).海馬の解剖学(3):海馬の神経解剖学(3). 前脳脊椎動物の共通プランの特徴。 魚類における空間的、時間的、感情的記憶のための特殊な口蓋領域。 Brain Res.ブル。 57: 397-399.
- Buzsáki, G (2002). 海馬におけるシータ振動. Neuron 33: 325-340.
- Buzsáki, G (2006). Rhythms of the Brain.
- Cantero, JL, Atienza M, Stickgold R, Kahana MJ, Madsen JR, Kocsis B (2003). ヒトの海馬と新皮質における睡眠依存性のシータ振動。 J. Neurosci. 23: 10897-10903.
- Colombo, M, Broadbent N (2000). 鳥類の海馬は哺乳類の海馬の機能的ホモログなのか? Neurosci. Biobehav. Rev. 24: 465-484.
- Eichnbaum, H; Cohen NJ (1993).
- Ekstrom, AD, Kahana MJ, Caplan JB, Fields TA, Isham EA, Newman EL, Fried I (2003).記憶、記憶喪失、海馬システム、MIT Press。 ヒトの空間ナビゲーションの基礎となる細胞ネットワーク。 ネイチャー425:184から188まで。
- Gray, JA; McNaughton N (2000年). 不安の神経心理学。 An Enquiry into the Functions of the Septo-Hippocampal System, Oxford University Press.
- Green, JD, Arduini AA (1954). 覚醒時の海馬の電気活動。 J. Neurophysiol. 17: 533-557.
- Jacobs, LF, Gaulin SJ, Sherry DF, Hoffman GE (1990). 空間認知の進化:空間行動の性特異的パターンが海馬の大きさを予測する。 PNAS 87: 6349-6352.
- Jacobs, LF (2003). 認知地図の進化. Brain Behav. Evol. 62: 128-139.
- Kirk, IJ, McNaughton N (1991). 乳頭上細胞発火と海馬の律動的緩慢活動。 Neuroreport 11: 723-725.
- Maguire, EA, Gadian DG, Johnsrude IS, Good CD, Ashburner J, Frackowiak RS, Frith CD (2000). タクシー運転手の海馬におけるナビゲーション関連の構造変化。 PNAS 97: 4398-4403.
- McNaughton, BL, Battaglia FP, Jensen O, Moser EI, Moser MB (2006). 経路統合と「認知地図」の神経基盤。 Nat. Rev. Neurosci. 7: 663-678.
- Moser, EI, Kropf E, Moser M-B (2008). 場所細胞、グリッド細胞、そして脳の空間表象システム。 Ann. Rev. Neurosci. 31.
- Nieuwenhuys, R (1982). 魚類の脳組織の概要. Am. Zool. 22: 287-310.
- O’Kane, G, Kensinger EA, Corkin S (2004). 深在性健忘症における意味学習の証拠。 患者H.M.との調査。 Hippocampus 14: 417-425.
- O’Keefe, J; Nadel L (1978). 海馬の認知地図, オックスフォード大学出版局.
- Portavella, M, Vargas JP, Torres B, Salas C (2002).海馬の認知地図, オックスフォード大学出版局. 金魚の空間学習、時間学習、情動学習に及ぼす終脳葉状病変の影響。 Brain Res.Bull. 57: 397-399.
- Rodríguez, F, Lópeza JC, Vargasa JP, Broglioa C, Gómeza Y, Salas C (2002).脳神経外科医が、金魚の空間学習、時間学習、情動学習に与える影響について検討した。 脊椎動物の進化を通じた空間記憶と海馬淡水域:爬虫類とテレスト魚類からの洞察. Brain Res.ブル。 57: 499-503.
- Sainsbury, RS, Heynen A, Montoya CP (1987). ラットにおける海馬タイプ2シータの行動的相関。 Physiol.Behav.誌 39: 513-519.
- Scoville, WB, Milner B (1957). 両側海馬損傷後の最近の記憶の喪失。 J. Neurol. Neurosurg. 20: 11-21.
- Shettleworth, SJ (2003). 餌を蓄える鳥類における記憶と海馬の特殊化: 比較認知の研究への挑戦. Brain Behav. 62: 108-116.
- Skaggs, WE, McNaughton BL, Wilson MA, Barnes CA (1996). 海馬の神経細胞集団におけるシータ位相の歳差運動と時間的シーケンスの圧縮。 海馬 6: 149-176.
- Squire, LR (1992). 記憶と海馬:ラット、サル、ヒトでの知見からの総合的な考察。 Psych. 99: 195-231.
- Squire, LR; Schacter DL (2002). The Neuropsychology of Memory, Guilford Press.
- Vanderwolf, CH (1969).「記憶の神経心理学」(日本評論社). ラットにおける海馬の電気活動と随意運動。 EEG & Clin. Neurophysiol. 26: 407-418.
- Vargas, JP, Bingman VP, Portavella M, López JC (2006).海馬の電気活動と随意運動. 金魚の終脳と幾何学的空間。 Eur. J. Neurosci. 24: 2870-2878.
- West, MJ (1990). 海馬の立体研究:ハリネズミ、実験用げっ歯類、野生マウス、人間を含む多様な種の海馬小区画の比較. Prog. Brain Res. 83: 13-36.
論文
- Amaral DGとCowan WM. 1980. サルの海馬形成に対する皮質下求心性神経。 ジャーナル・オブ・コンパラティブ・ニューロロジー。 2月15日; 189(4):573-91.
- Duvernoy, H. (2005) The Human Hippocampus, 3rd ed.ベルリン: Springer-Verlag.
追加資料
論文
- Google Scholar
- http://www.psycheducation.org/emotion/hippocampus.htm Great brain tours!!!!
- BrainMaps at UCDavis hippocampus
- Artificial Hippocampus
- BrainInfo at the University of Washington hier-164
- John O’Keefe & Lynn Nadel (1978) The Hippocampus as a Cognitive Map 、Oxford University Press. Full Text
Additional images
このギャラリーに写真を追加する
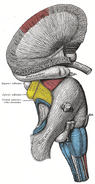
ヒトの脳。 大脳辺縁系
扁桃体 – 帯状回 – 叉状回 – 海馬 – 視床下部 – 乳頭体 – 側坐核 – 眼窩前頭皮質 – 海馬傍回
|
v-d-e
人間の脳です。 前脳(大脳-大脳皮質-大脳半球、灰白質)(TA A14.1.09.002-240, 301-320, ga 9.818-826) |
||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 前頭葉 |
|
|||||||||||||||
| 頭頂葉 |
|
Superolateral
|
内/下
|
両方
|
||||||||||||
| 後頭葉 |
|
上外側
|
内側/内側
|
|||||||||||||
| 側頭葉 |
|
上外側
|
内側/下
|
|||||||||||||
| 葉間 溝・裂 |
の3つ。 |
超外側
|
内側/下
|
|||||||||||||
| 大脳辺縁系 |
|
その他
|
||||||||||||||
| 島状葉
島嶼長回 – 島嶼短回 – 島嶼円形溝 |
||||||||||||||||
| 一般 |
大脳皮質 – 大脳半球の極 |
|||||||||||||||
いくつかの分類は概算である。 また、一部のブロドマン領域はジャイロにまたがっている。
このページはWikipedia(作者表示)からのクリエイティブ・コモンズ・ライセンスを使用するコンテンツが含まれています。
|
||||||||||||||||