解糖の後半(エネルギー放出ステップ)
これまで、解糖は細胞に2つのATP分子のコストをかけ、小さな炭素3糖分子を2つ作り出しました。 これらの分子は両方とも経路の後半を進み、初期投資として使われた2つのATP分子を回収し、さらに2つのATP分子とさらに高エネルギーのNADH分子2つという利益を細胞にもたらすのに十分なエネルギーが抽出されるであろう。 解糖の第6段階(図9.1.2)では、糖(グリセルアルデヒド-3-リン酸)を酸化して高エネルギーの電子を取り出し、それを電子伝達物質NAD+が拾ってNADHを生成する。 その後、糖は2つ目のリン酸基を付加されてリン酸化され、1,3-ビスホスホグリセリドが生成される。 2つ目のリン酸基は別のATP分子を必要としないことに注意。
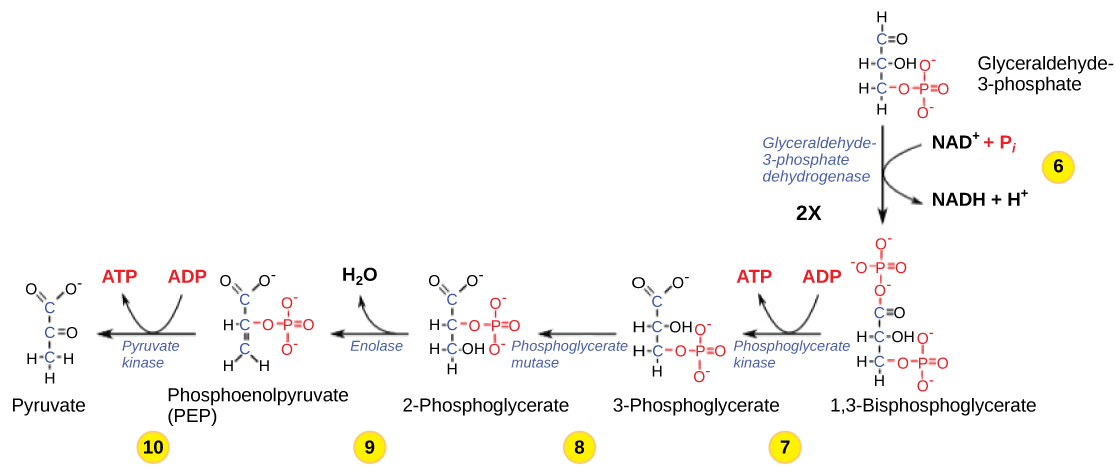
ここで再びこの経路の制限要因となりうるものが出てきた。 反応の継続は、電子キャリアーであるNAD+の酸化型の利用可能性に依存する。 したがって、この段階を継続させるためには、NADHが継続的に酸化されてNAD+に戻らなければならない。 NAD+が利用できない場合、解糖の後半は遅くなるか停止する。 もし系内に酸素があれば、NADHは間接的ではあるが容易に酸化され、この過程で放出される水素からの高エネルギー電子がATPの生成に利用されることになる。 酸素のない環境では、代替経路(発酵)によりNADHからNAD+への酸化を行うことができる。
ステップ7.NADHからNAD+への酸化は、NADHからNAD+への酸化である。 第7ステップでは、ホスホグリセリン酸キナーゼ(逆反応のために名付けられた酵素)が触媒となって、1,3-ビスホスホグリセリンが高エネルギーのリン酸をADPに供与し、ATP1分子を形成する。 (1,3-ビスホスホグリセリン酸のカルボニル基はカルボキシル基に酸化され、3-ホスホグリセリン酸が生成する。 第8ステップでは、3-ホスホグリセレートに残ったリン酸基が第3の炭素から第2の炭素に移動し、2-ホスホグリセレート(3-ホスホグリセレートの異性体)が生成される。 このステップを触媒する酵素は、ムターゼ(異性化酵素)である
ステップ9. エノラーゼが9番目のステップを触媒する。 この酵素は2-ホスホグリセリドの構造から水を失わせる。これは脱水反応であり、その結果、二重結合が形成されて、残りのリン酸結合の位置エネルギーが増加し、ホスホエノールピルビン酸(PEP)が生成される。 解糖の最後のステップは、ピルビン酸キナーゼという酵素によって触媒され(この場合の酵素は、ピルビン酸のPEPへの変換の逆反応から名付けられた)、基質レベルのリン酸化とピルビン酸(またはその塩形態、ピルビン酸)という化合物によって第2のATP分子が生成されることになる。 酵素経路の多くの酵素は、酵素が順反応と逆反応の両方を触媒できるため、逆反応の名前が付けられている(これらは当初、非生理的条件下、試験管内で行われる逆反応によって説明されていたのかもしれない)。
グルコースのピルビン酸への変換における純反応は、以下のとおりである:

したがって、2分子のATPが、グルコースの2分子のピルビン酸への変換で発生させられることになる。
なお、グルコースの2分子のピルビン酸への嫌気性変換で放出されるエネルギーは、-21kcal mol-1(-88kJ mol-1)である。