Introduction
生態学者や生物地理学者は、陸上植物の多様性を気候的に決まるバイオームに整理し、支配種の成長形態や機能形質によって人相を特徴とする(Moncrieff et al, 2016). 実際、バイオームの概念は、カール・ルードヴィヒ・ウィルデナウとアレクサンダー・フォン・フンボルトまでさかのぼる長い歴史を持っている。 ウィルデナウは、類似した気候が類似した植生形態を支えていることを認識し、フンボルトは、植物の分布、人相、環境要因の間に広範な関連性があることを観察した。 バイオームの概念は、進化の歴史とは無関係に、同じような環境圧力が同じような機能的属性を持つ種を選択する、という仮定を反映している。 同時に、地球上の現存するバイオームは、ある程度系統的に区別されており、特徴的な種の多く/大部分は、気候の選択だけでなく、火災や巨大草食動物などの追加圧力に対する重要な適応を示す特定の系統から引き出されている(Woodwardら、2004;Penningtonら、2006; Donoghue and Edwards, 2014)。 バイオームは地球の植生における広範な規則性を表すため、バイオーム間の機能の違いを理解することは、地球規模の炭素循環と地球システムの機能(人為的な地球変動に対する反応を含む)をモデル化する上で決定的に重要である(Bonan et al, 2012; van Bodegom et al., 2014; Xia et al., 2015)。
過去20年間、大陸スケールにわたる植物生物多様性の機能的次元の理解が急速に進んでいる(Swenson et al., 2012; Lamanna et al., 2014; Šímová et al., 2018)。 広範なデータ合成と分析の努力により、機能的形質スペクトルとして知られる、植物全体のサイズ、葉の経済性、および水力学の変化を組み込んだ限られた機能的形質空間が定義されている(Wrightら、2004;Díazら、2016)。 機能形質スペクトルは、植物の生活史戦略を決定する生理的・生態的トレードオフを定義し(Westoby, 1998; Reich et al., 2003; Craine, 2009)、群集形成(Kraft et al., 2008)、生態系機能(Lavorel and Garnier, 2002; Garnier et al., 2004; Kerkhoff et al., 2005; Cornwell et al., 2008)、さらには進化率(Smith and Donoghue, 2008; Smith and Beaulieu, 2009)に影響を及ぼすと考えられている。 植物機能形質スペクトルの汎用性と有用性は、植生の地球規模の分布(van Bodegomら、2014)、生物地球化学循環(Bonanら、2012)、生態系サービス(Díazら、2007、Cadotteら、2011、Lavorelら、2011)のモデルへの取り込みを急がせている。 しかし、支配的な種と従属的な種や生命体の両方について、バイオームスケールで機能的多様性の包括的な尺度を調べた研究はほとんどない。
気候的・人為的に異なるバイオーム間で機能的多様性がどのように異なるかについては、対照的な2つの予測セットが存在する。 一方では、バイオメスの生物多様性が利用可能な生態学的ニッチ空間の変動を反映している場合、分類学的に多様でないバイオメスは、より多様なバイオメスが占める機能空間のより小さい、大きく入れ子状のサブセットを表すはずである。 一方、植物の構造と機能における基本的なトレードオフのグローバルな性質と、種間で同様に作用する同様の選択圧により、異なる環境を占める群体は、実際には形質空間の同様の領域を共有しているかもしれない(Reichら、2003;Wrightら、2004;Díazら、2016)。 最近の研究では、気候と機能的・分類学的多様性(すなわち、種の豊かさ)との関係は複雑であり、規模に依存することが確認されている。 場合によっては、機能的多様性は気候勾配を密接に追跡し、より変動的で極端な条件では機能的多様性が低くなり(Swenson et al., 2012; de la Riva et al., 2018)、局所的な集合体における機能空間の容積は、分類学的豊かさの増加とともに拡張する(Lamanna et al., 2014; Li et al., 2018)。 しかし,地域規模の種プールの分析から,種の豊かさの大きな変化は,機能的多様性への最小限の影響と関連している可能性が示唆されている(Lamanna et al., 2014; Šímová et al., 2015)。 さらに、気候に対する分類学的及び機能的多様性の反応は、主要な植物成長形態間で異なる可能性があり(Šímová et al., 2018)、異なる成長形態の相対的多様性は、異なるバイオームを定義する気候勾配に沿って大幅に変化するかもしれない(Engageann et al., 2016)
大陸から地球スケールでの機能的多様性の分析の進展は、一部、生物多様性の分類学、分布、系統及び機能の側面を目録するデータ不足のために限られている(Hortal et al., 2015)。 さらに、利用可能なデータが限られている場合でも、種の地理的分布、機能的特性、系統的関係を記述する大規模で異質なデータセットを標準化し統合するためには、情報科学上の大きな課題がある (Violle et al., 2014)。 ここでは,陸上植物の>10万種に関する分布と機能形質の情報を集めたBotanical Information and Ecology Networkデータベース(BIEN; Enquist et al., 2016; Maitner et al., 2018)を用いて,北米と南米のバイオマスにおける植物種の機能多様性の分布を調べる。 具体的には、100×100kmグリッドセル規模の陸上植物種分布図と、生態戦略の変動の主要軸を反映する6つの機能形質の包括的なデータセットに基づいて、バイオームの機能的多様性と識別性の変動を検証する
本研究の目標は3つである。 まず、持続的なデータ不足を強調するために、バイオームの機能的多様性と独自性を特徴付ける利用可能なデータの範囲を文書化する。 第二に、利用可能なデータに基づいて、バイオーム内の機能的多様性の分布を、支配的な成長形態と従属的な成長形態の両方について明らかにすることである。 これらの分析により、バイオーム内で最も広く分布する種の最も一般的な機能戦略を特定することで、バイオームの機能的独自性をより適切に定量化することができる。 第三に、機能的形質空間における多次元超体積の重複に基づいて、バイオームが実際に機能的に異なる種のコレクションによって特徴付けられるかどうかを問う。
方法
分布データおよびバイオーム分類
複数のリソースからコンパイルした発生データセットに特有のサンプリングの偏りの影響を減らすために、北および南アメリカにおいて分布する植物種の88,417に対するBIEN 2.0 range maps(ゴールドスミスら, 2016). BIENデータベースは、植物標本と植生プロットインベントリに由来する標準化された植物観察結果を統合したものである。 BIENデータベースの種の範囲マップは、それぞれの種について利用可能な発生数に応じて、3つのアプローチのうちの1つを使用して推定された。 出現数が1つか2つしかない種(種の約35%)については、地理的範囲を各データポイントを囲む75,000km2の正方形のエリアとして定義した。 3〜4回出現する種の地理的範囲は、凸包を使用して特定した(約15%の種)。 最後に、少なくとも5つの出現を持つ種の範囲マップは、モデルによる過剰な予測を抑制するために、19の気候層を予測変数として、19の空間固有ベクトルをフィルターとして使用するMaxent種分布モデリングアルゴリズムを用いて得られた(Goldsmith et al, 2016 for further details on range map methodology)。
ランバート方位等面積投影による100×100kmグリッドマップにBIEN 2.0植物種マップを重ね、各グリッドセルに対する種の存在/不在マトリックスを得た。 Olson et al. (2001)のバイオーム分類に基づき、各マトリックスグリッドセルをバイオーム分類の一つに割り当てた。 その後の分析には計算上の限界があるため、気候や植生、文献の類似性に基づいていくつかのバイオームを結合し、表1に示すような大分類とした(内陸水域、岩石と氷、マングローブを除く)。 チャコとカアティンガのエコリージョンは,Prado and Gibbs (1993), Pennington et al. (2000), Banda et al. (2016), Silva de Miranda et al. (2018) に従ってそれぞれXeric Woodlands, Dry forestに分類した。
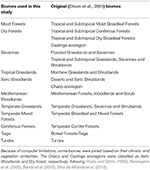
Table 1.表1.のとおりである。 本研究で採用したバイオーム分類と同等の(Olson et al., 2001)バイオーム分類の概要(Inland Water, Rock and Ice, Mangrovesを除く)
Species Composition Among Biomes
バイオス間の機能形質空間の変動を理解するために、まずバイオス間の植物種構成の差異を探った。 各グリッドセルとバイオームの種リストに基づき、各バイオームの特徴的な種を、そのバイオームでその範囲の最大割合を占める種として定義した。 これらの、あるバイオームで地理的に優勢な種のリストを、他のバイオームの種のリストと比較した。 この一対の比較は、生物群間の方向性のある分類学的重複(すなわち、ある生物群の優勢な種が他の生物群と共有する割合)を推測する簡単な方法を提供するものである。 範囲マップが利用可能な88,417種のうち、44,899種は複数のバイオームにまたがる範囲を持ち、43,518種は特定のバイオームに固有である。
特性データ
我々はBIEN 3.0 dataset(2018年2月7日に取得)で利用できる新世界の植物種のすべての特性情報を抽出し、合計80405種レベルの特性観測を得た。 次に、最大草丈(m)、種子質量(mg)、木材密度(mg/cm3)、比葉面積SLA(cm2/g)、単位質量あたりの葉のリンと葉の窒素濃度(mg/g)という6つの機能形質について情報をフィルターにかけた。 フィルタリングの結果、合計 18,192 件の種レベルの観測値が残った。
各バイオームの機能形質空間を推定するためには、完全な形質データが必要であった。 このため,Rパッケージ「Rphylopars」v 0.2.9 (Goolsby et al., 2017) とSmith and Brown (2018) が最近発表した種子植物の系統樹を基準として,欠損形質データを系統樹的にインプットした (Bruggeman et al., 2009; Penone et al., 2014; Swenson, 2014; Swenson et al., 2017). 系統的インピュテーションは、近縁種は類似の形質値を持つ傾向があるという仮定に基づき、機能的形質データセットにおける欠損データを予測するためのツールである(Swenson, 2014)。 我々は、ALLBM木(すなわち、Magallónら、2015によって提供されたバックボーンを有する遺伝子バンクおよびOpen Tree of lifeの分類群)を使用したが、それは利用可能な形質および分布情報を有する種間の重複を最大化するものであったからである。 形質インピュテーション後、完全な形質情報と範囲地図を持つ合計7,842種がさらなる分析のために残った。
形質ハイパーボリューム測定
我々は、バイオメスセル内の種プールから形質ハイパーボリュームを計算することによって、バイオメスの相対機能多様性を測った。 計算機の制限により、各バイオームのセルの20%のランダムサンプルを用いて形質ハイパーボリュームを構築した。 各グリッドセルのハイパーボリュームは,抽出した6つの機能形質とRパッケージ「hypervolume」(Blonderら,2014,2018)を用いて,デフォルトのSilverman帯域幅推定器を用いたガウスKDE法により推定した。 種子質量、高さ、木材密度は対数変換され、一方、SLAは平方根変換された。 すべての形質は解析前にスケーリングおよびセンタリングされた。 ハイパーボリュームは標準偏差(sd)の単位で、使用した形質の数に対するべき乗(すなわち、sd6)で報告する。
機能的独自性と広域性
バイオームはその優勢な植生によって特徴づけられるため、バイオーム内の種の地理的共通性と機能的独自性についても検討した。 種の範囲地図と機能形質情報を用いて,Violleら(2017)による機能的希少性の概念的枠組みに従い,各バイオームにおける各種の機能的独自性と広範性を推定した。 測定およびインプットされた)形質の全セットを用いて、まず、すべてのペアの種間の標準化された形質空間におけるユークリッド距離を測定した。 次に、各バイオームにおける各生物種の機能的識別性(Di)を、バイオーム種プール内のN個の他の種に対する種の平均機能距離として算出した。 Diは、バイオーム内の他の種と比較して、低い値は機能的に共通(冗長)である種を表し、高い値は機能的に特徴的である種を表し、各バイオーム内で0と1の間でスケーリングされた。 また、バイオーム内の種の地理的な広さ(Wi)を推定し、バイオーム内の全セル数に対する種が占めるグリッドセルの数として測定した。 1の値は、そのバイオームがカバーするすべてのグリッドセルに対象種が存在することを示す。 機能的識別性と地理的広域性の両方をRパッケージ「funrar」(Grenié et al., 2017)を用いて算出した。
局所的な存在量またはバイオマスに基づく優越性の包括的指標がないため,各バイオームについて,地理的に広域で(Wi > 0.5) かつ機能的に類似した(Di < 0.25)として最も「ありふれた」種を識別するのに指標として Di および Wi を使用した. バイオーム間の機能的差異値の第3分位が0.2〜0.3であったため、この最後の閾値が用いられた。 機能的に異なる種と機能的に類似した種をその後の分析で区別するために、0.25をカットオフ値として使用した(すなわち。
機能空間とバイオームの類似性
バイオームの超体積間の重複を推定するために、(i) 種の総数、(ii) 各バイオームで機能的に共通で地理的に広いと考えられる種のリストを使ってSørensen類似性指標を採用した。 機能的形質空間の類似度は、生物群間のハイパーボリュームのペアワイズ分数オーバーラップとして算出した。 分数的重複は,2つの超体積の交点の体積の2倍を,それらの結合部の体積で割ることによって計算された. すべてのハイパーボリュームは,R パッケージ “hypervolume” を用いて推定した(Blonder et al., 2014, 2018)、デフォルトのシルバーマン帯域幅推定器を用いたガウスKDE法を使用した。
結果
我々は、モデル化した種の範囲の交差に基づいて、バイオマス間で植物種の組成にかなりの重複を発見した(図1、および補足表1参照)。 この分類学的重複は熱帯と温帯のバイオーム内でより大きく、これら2つの気候帯間で共有される種は比較的少なかった。 興味深いことに、乾性森林地帯は熱帯と温帯の両方のバイオームと種を共有している。 熱帯湿潤林に特徴的な種の割合が高い(83%)にもかかわらず、このバイオームは乾燥林、サバンナ、乾性森林、熱帯草原など他の熱帯バイオームとも多数の種を共有している。 温帯草原、地中海森林地帯、タイガ、ツンドラなどの温帯バイオームの高い分類学的重複は、いくつかの温帯バイオームが、より種が豊富で機能的に多様な熱帯バイオームの劣ったサブセットを代表して、低い機能分化の可能性を示唆している
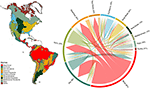
Figure 1. 新世界のバイオーム間の植物種の重複。 パーセンテージの値は、そのバイオーム(N = 84,413)に地理的範囲の最大の割合を持つバイオームに発生する種の割合を表す。 各分岐矢印の基部は、種の範囲の最大の割合を含むバイオームを示すように配置され、幅は矢印の先端にあるバイオームと共有する種の数を表す。
マッピングされた種の10%未満しか表現されていないため、我々のバイオームスケールの種群に対して形質データはかなりまばらであった(図2A)。 さらに,利用可能な形質データは,形質,植物クレード,バイオーム間で大きく異なっていた. 系統学的に見ると、葉身と木密は特に表現が乏しく、利用可能なデータが全種類に欠けていた。 その結果、7,842種の形質値の約2/3を帰属させる必要があったが、その多くは近縁種が存在しないために帰属させなければならなかった。 地理的には、SLA、樹高、種子量は熱帯と温帯の南米で十分に抽出されていないが、葉のNと木材密度は地域間でより均等に抽出されている。 葉身はすべてのバイオームでサンプル数が少ない。 各バイオームの20%のセルからランダムに選択した種プールから作成した形質ハイパーボリュームは、主に熱帯と温帯/寒帯のバイオームの間で違いが見られる(図2B)。 形質ハイパーボリュームは熱帯バイオームでより大きく、より変動する傾向があり、湿潤熱帯林と乾燥熱帯林で最も変動が大きい。 温帯/寒帯バイオームでは、針葉樹林と温帯混合林が最も大きな形質超体積を支えている。 興味深いことに、乾性低木林は温帯バイオマスよりも熱帯バイオマスに近い過体積の分布を示す。
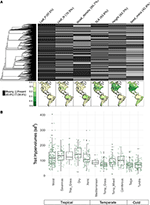
Figure 2.2.に示すように、熱帯バイオマスの過体積の分布は温帯バイオマスよりも熱帯に近い。 (A) BIEN 3.0データベースにおける、既知(灰色)または欠損(黒色)の形質値を持つ種の割合。 左の系統図は、Smith and Brown(2018)の種子植物ALLBMツリー(すなわち、Magallónら、2015によって提供されたバックボーンを持つGenBankおよびOpen Tree of life分類群)に相当する。 マップは、BIEN 2.0データベース内の種の総数に対する、形質情報を持つ種の割合を表す。 (B)各生物群の100×100kmのセルからランダムに選択した20%の形質ハイパーボリュームの分布。 ハイパーボリュームは使用した形質数の標準偏差のべき乗の単位で報告されている。
機能的識別性と地理的分布の関係はバイオーム間で驚くほど類似している(図3)。 どのバイオームでも、大多数の種は地理的に制限され、かつ機能的に類似している。 同時に、すべてのバイオーム内で最も機能的に特徴的な種は、一般的に地理的にも制限されており、最も地理的に広く分布する種は、ほぼ常に機能的に類似していた
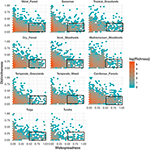
Figure 3. バイオーム全域における機能的差異性のパターン。 識別性は,バイオーム内で種が互いにどの程度機能的に離れているかを表す(すなわち,焦点となる種から他のすべての種までのペアワイズ表現型距離の平均値). この値が大きいほど、その生物群の機能空間のセントロイドから遠い種である。 広域性(Widespreadness)は、ある種がバイオーム内で地理的にどの程度一般的であるかを測定する。 値が 0 の場合、その種が 1 つのバイ オームセルに存在することを示す。 Bin 値は対数スケールでの種の数である。 長方形は、各バイオームにおいて機能的に類似し、広く分布していると考えられる種を、それぞれ区別性と広範性のカットオフ値0.25と0.5を用いて表した。
我々は、バイオーム内およびバイオーム間の成長形態の比例分布にかなりの変動を見出した。 熱帯のバイオームでは木本類が種数の点で支配的な成長形態であるのに対し、温帯の環境では草本類が支配的である(図4)。 各バイオームで広く分布し、機能的に共通する種のみを考慮した場合、バイオーム間の成長形態の分布は変化し、特に樹木、草本、草の割合が大きくなった。 例えば、熱帯のバイオームでは、湿潤林を除く各バイオームで樹木の割合が減少した。 温帯のバイオームでは、特に極地に向かって草の割合が増加した。 興味深いことに、広く分布し、機能的に一般的な種のみを考慮した場合、木質林における成長形態の分布は温帯バイオームにおける分布により近い。
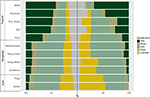
Figure 4. 各バイオームにおける成長形態の分布(左)は種の総数を用い、(右)は機能的に類似し、バイオーム内に広く分布する種のみを用いた(図3参照)
バイオーム間の種構成の一対比較は、熱帯、温帯、極地の気候帯を表す3つの主要なクラスターを明らかにし(図5A)、これらの地域間ではなく、その中で高い分類学の重複を反映している(図1)。 バイオマス間の形質超体積の一対比較では、気候帯のクラスタリングはそれほど明確ではないことが示された(図5B)。 この場合、乾性森林地帯の機能空間は温帯バイオームと大きく重なっている。 この分析により、タイガの機能空間が温帯草原や混合林と重複していることも明らかになった。 機能的に共通で広く分布していると考えられる種のみを用いたバイオマス間の形質超大の一対比較では、気候帯間および気候帯内の形質空間の重複が少ないことが示された(図5C)。 しかし,現在では,木質森林地帯が他の熱帯バイオームとクラスタリングされているにもかかわらず,これらの生息地は熱帯草原とともに,地中海森林地帯や温帯草原などの温帯バイオームと機能空間において大きな重複を示す
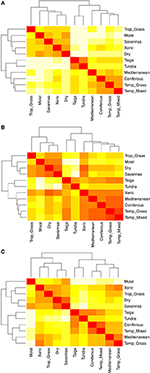
FIGURE 5. (A)バイオマス間の種組成のペア非類似度。 (B)種の総数を用いたバイオマス間の形質超大型化(1-Sørensen類似度)のペアワイズ非類似度. (C)機能的に類似し、かつ広く分布していると考えられる種のみを用いたバイオマス間の形質超体積のペアワイズ非類似度(1-Sørensen類似度)。
議論
我々の分析は、機能的なレンズを通して陸上バイオームを理解するための3つの重要な洞察をもたらす。 第一に,生物多様性の一次データの編集と統合が進んでいるにもかかわらず,大陸から地球規模まで生物群の機能的生物多様性を定量化する我々の能力を制限しかねない重大な知識不足が存在することを示す。 第二に、入手可能なデータの分析から、南北アメリカのすべてのバイオームには、地理的に最も広く分布する種が共通の機能特性を持つ一方で、最も機能的に特徴的な種は常に分布が限定されるという、驚くべき共通パターンがあることがわかった。 第三に、広く分布し、機能的に共通する種のみを考慮した場合、バイオームをより容易に機能的に区別することができ、バイオーム間の非類似パターンは、気候と植物機能ニッチ空間の対応を反映しているように思われる。 まとめると、我々の結果は、バイオームの機能的多様性の研究はまだ形成段階にあるが、この分野のさらなる発展により、進化、生物地理学、コミュニティアセンブリ、および生態系機能を関連付ける洞察が得られる可能性が高いことを示唆している。 例えば、マッピングされた種のうち、形質データと系統情報の両方が利用可能なものは10%未満であっただけでなく、それらの欠落した形質データ(種の形質値の〜66%)は、系統的にも地理的にもかなり構造化されていた。 このように、機能的形質データにおけるラウンキア的不足(sensu Hortal et al., 2015)は、分布情報におけるウォーレス的不足、系統的理解におけるダーウィン的不足と相互作用しているのです。 そして、それらの欠損値を合理的に埋め込むために系統的知識を用いることができるが(Swenson et al.、2017)、バイオーム分類にとって重要な結果のパターンのいくつかの側面は依然として不確実である。 例えば、既存の成長形態データは、すべての熱帯バイオームで木本種が優勢であることを示唆しているが、温帯および極地バイオームでは草本の成長形態の割合的多様性がはるかに高い(Engemannら、2016;Zanneら、2018も参照のこと)。 草本植物の相対的多様性におけるこの緯度方向の増加は,サンプリングバイアスや熱帯の草本植物に関する分類学的知識の欠如を反映しているのか,それとも近極と新熱帯の領域における進化的・生物地理学的歴史の違いを反映しているのか? 支配的な成長形態はバイオームを区別する重要な特徴の 1 つであるため、最も多様な成長形態における系統的な変化(支配的かどうかにかかわらず)は、バイオーム全体で機能的多様性がどのように変化し、その変化が生態系の機能とサービスにどのように影響するかについての予測に必ず影響する。 しかし、種の分布モデルから描かれた範囲マップは、我々の結果における不確実性の追加要素を表す可能性がある。 例えば、本研究の地理的解像度と特定のバイオームの空間的複雑性を考慮すると、これらのモデルは、単一のバイオームのセルから近接・相互接続したバイオームのセルまで、いくつかの種の地理的範囲を過大評価する可能性がある。 しかし、バイオーム間の機能的重複がわずかに減少することで、いくつかの種の範囲が実際に過大評価されていることが明らかになった(補足図2、5、6を参照)。 また、温帯と熱帯の相対的なオーバーサンプリングにより、出現情報を使用することで、我々の結果における機能的独自性と広域性の一般的な関係も変化した(補足図4参照)。 この場合、熱帯バイオームにおけるほとんどの種のサンプリングが極めて限られているため、我々の広範性の尺度は制限されることになる。 データセットとサンプリングが改善されれば、将来的にこれらの不確実性を減らす余地がありそうだ。
バイオメスの機能的多様性を理解するために取り組むべき他のデータギャップは、存在量情報のPrestonian不足である。 群集形成、生態系機能、生物地球化学的循環にとって最も重要な形質は最も支配的な種のものであるため、存在量は機能的多様性の研究において特に重要である。 我々は、種分布モデルから得られた範囲マップに基づいて集合体を作成したため、共通性の地理的要素にしか対処できなかったが、局所的な存在量と地理的範囲の間の正の関係、いわゆる占有-存在量関係は、各バイオームで最も普及している種は、しばしば、より豊富でもあることを示唆している(Borregaard and Rahbek, 2010; Novosolov et al, 2017)。 バイオームは一般的にその地域で支配的な生育形態によって特徴付けられるため、豊度情報を統合することで、機能的な重複が少ないバイオームを実現できるかもしれない。 しかし、BIENが地域の存在量を推定できる膨大なプロットデータを編集しているにもかかわらず、異なる地域や成長形態間で妥当な比較を行うことは、不均一なサンプリング、ガンマ多様性の地域差、存在量を定量化する非整合な方法によって妨げられています。
Common Patterns Within Biomes
利用可能なデータのギャップがあるにもかかわらず、我々の分析は、西半球のすべての生物群に共通する、これまで記録されていない関係を明らかにした。 この「占有率-冗長性」の関係は、バイオーム内の気候条件が一連の共通特性を選択し、より機能的に特徴的な種はバイオーム内の生息地の希少な部分集合に限定されることを示唆しているのかもしれない。 すべてのバイオームで機能的に類似した種が全体的に多いことと、「占有率-冗長性」の関係は、生息地のフィルタリングが機能的に類似した種の共存につながることを示唆するコミュニティレベルの機能多様性に関する最近のグローバルな分析(Li et al.、2018)や、機能の冗長性がコミュニティの安定性を高めることを示す研究(例えば、Walker et al.、 1999; Pillar et al.、 2013)と一致しています。 さらに、バイオーム間で分類学的および機能的な重複が高いこと(図1、5)とともに、一般的に広く分布する種が機能的に類似しているという事実は、幅広い環境条件にわたる陸上植物が、機能特性スペクトルの中核付近に共通の特性を有しているという考え方を補強する(ライトら、2004;ディアズら、2016)。 Umañaら(2017)とは異なり、我々の結果は、希少で地理的に制限された種が、より広く一般的な種と機能的に異なる可能性があること、またはそうでない可能性があることを示すものである。 したがって、より機能的に特徴的な種が特定の環境における専門家なのか、あるいはその形質の組み合わせが、地理的分布を制限する人口学的または生理学的トレードオフをもたらすのか、疑問が残る。
生物群間の比較
生物群間の形質超体積分布(図2B)を比較すると、より種の多い環境もより機能的に多様という観測と一致する(Swenson et al)。 2012; Lamanna et al., 2014; Li et al., 2018; Šímová et al., 2018)と一致する。 我々の結果は、季節的・極端な気候環境が一貫して種の機能的多様性を制限するという仮説(de la Riva et al.、2018)に関しては、より不明確である。 すべての熱帯バイオームがすべての温帯バイオームよりも高い平均機能多様性を示し、極地バイオームは最小のハイパーボリュームを示す。 しかし,各グループの中で,乾燥したまたは季節変動の大きいバイオームは,Currieら(2004)による耐性仮説に従って予想されるように,必ずしも小さいハイパーボリュームを表示しない(例えば,乾燥林,乾性森林),一方,最適な気候条件は,生理学的パラメータのより多くの組み合わせをサポートする。 これらの結果だけでは、温帯・極地バイオームが機能的ニッチ空間の制限により分類学的多様性が低いのか、それとも分類学的多様性が低いために機能的ハイパーボリュームが小さいのかを判断することはできない。 ヌルモデリングアプローチは,分類学的多様性と機能的多様性を切り分けるのに役立つ可能性があるが(Swenson et al., 2012; Lamanna et al., 2014; Šímová et al., 2018),そうした分析はこの研究の範囲外だった。 より重要なのは、我々の結果が、進化と生物地理学の歴史がバイオームの機能的多様性をどのように形成するかを理解することの重要性を補強することである(Woodwardら、2004;Penningtonら、2006;Donoghue and Edwards、2014;Moncrieffら、2016)。 異なる生物地理学的領域間ではなく、領域内の生物界をまたぐ種(および高等系統)の広範な共有は、領域内の機能的多様性の均質化と、異なる環境によって選択される、あるいは選択されない特徴的形質に関する手がかりの両方に役立つと考えられる(ドゥーマら、, 2012; Zanne et al., 2014, 2018)。
すべてのバイオーム間でかなりの超体積重複があるにもかかわらず(補足図2)、熱帯、温帯、および寒冷バイオームはすべて機能空間の区別できる領域を占めているようだ(図5B,C)。 バイオームを区別する主な形質は、より局所的なプロットベースの分析(例えば、Douma et al.2012)で観察されたような葉の経済的形質ではなく、成熟した高さと種子量の両方を含む植物全体のサイズに関する形質(補足図7-10)であるように思われる。 例外は葉のPで、熱帯と温帯・極地のバイオーム間でかなりの差が見られた(補足図7-10)。このパターンは、種固有の値と生態系全体の測定値に基づく他の分析でも観察されている(Kerkhoff et al.、2005;Swenson et al.、2012)。 葉身は最もサンプル数の少ない形質であるため、熱帯と温帯・極地の間の差は、インピュテーションの手順によるバイアスに左右される可能性がある。 しかし、葉のPは有意な系統的シグナルを示し(Kerkhoff et al.、2006)、これはインピュテーションがバイアスを受けないことを示唆している(Swenson et al.、2017)。 葉のPは草本の成長形態でも有意に高い(Kerkhoff et al., 2006)ため,木本から草本への優勢な多様性(図4)への緯度シフトもこのパターンに影響を与えるかもしれない。
超体積分析を各バイオームで最も広く,機能的に共通の種のみに限定すると,各生物地理的領域内の個々のバイオームは機能特性空間において重複が少なく(比較図5B,C),これらの種は特定の環境に適応した表現型が反映していると思われる. この中で、特に興味深いのは、木質森林バイオームである。 チャコエコリージョンが含まれていることもあり、分類学的にはきせい森林バイオームが熱帯バイオームと一緒になっている(図5A)。 しかし、すべての種の機能的なハイパーボリュームを考慮すると、温帯バイオームとはるかに強い類似性を示す(図5B)。 最後に、最も広く分布し、機能的に共通する種のみを分析した場合、木質林は再び熱帯バイオスとの類似性が高まるが、温帯草原や地中海バイオスとの高い類似性も維持される(図5C)。 温暖な中生環境から、より寒冷で乾燥した、より季節感のある環境への移行は、類似した形質、例えば草本習性によって促進され(Douma et al., 2012; Zanne et al., 2014, 2018)、寒冷環境同様、乾燥環境の群集も系統的にクラスター化する傾向にある(Qian and Sandel, 2017)。 さらに、熱帯収束帯の境界に位置するキセリックゾーンは、熱帯域と温帯域の間の地理的な移行点となる。 xeric woodlandsが機能的にも生物地理的にも熱帯域と温帯域の中間であるという事実は、バイオメスの機能的多様性をよりよく理解するためには、その生物地理的・系統的歴史を考慮しなければならないという考えをさらに強固にする(Pennington et al.2006; Donoghue and Edwards, 2014; Moncrieff et al…)。 2016)。
結論
陸上バイオームのいかなる分類も、気候や種分布の連続勾配に少数の離散的カテゴリーを押し付け、したがって複雑な生態学的景観の総簡略化を意味する(Moncrieffら、2016、しかしSilva de Mirandaら、2018を参照のこと)。 しかし、過度の単純化の可能性にもかかわらず、バイオームは地球の主要な陸上生態系の生物多様性と機能を整理し理解するための有用な構成要素であり、形質ベースのアプローチは世界の植生分布の動的モデル化に役立つ高い可能性を持つ。
本研究では、すべてのバイオームの分類学的多様性の多くが狭い分布と機能的に同様の種を示していることが示された。 さらに、各バイオーム内で最も機能的に特徴的な種は地理的に希少である傾向があり、一方、広く分布する種は一様に低い機能的特徴しか示さない。 生物群間における分類学的・機能的重複が大きいにもかかわらず、生物群は、特に各生物群に最も広く分布し機能的に共通する種に基づくと、生物地理的・気候的に区別可能な単位に集積することができる。 しかし、バイオームの機能的理解を進めるには、植生タイプ内および植生タイプ間の形質変異のより良い特性化だけでなく、種群の生物地理学的および系統発生史、バイオス内の種の相対存在度に関する情報も必要となる。
データ入手
本研究で分析した形質データセットおよび種範囲マップはRパッケージBIEN (Maitner et al., 2018) を通じてダウンロードできる。 BIENデータベースの詳細についてはhttp://bien.nceas.ucsb.edu/bien/を参照
著者貢献
SE-L、AK、BJE、DMN、CVは研究を設計し、SE-LとAKはデータを分析し、SE-LとAKは原稿の最初のバージョンを書き、CV、BB、NK、BSM、BM、RKP、BS、SS、J-CS、SKWが、特質と範囲地図の照合およびデータベース作成に貢献した。 7817>
Funding
SE-L, DMN, AKは米国国立科学財団の共同研究助成金(DEB-1556651)の支援を受けている。 BJEはNational Science Foundation award DEB-1457812およびMacrosystems-1065861の支援を受けた。 CVは、CESABプロジェクトCauses and consequences of functional rarity from local to global scalesの文脈で、欧州研究評議会(グラントERC-StG-2014-639706-CONSTRAINTS)とフランス生物多様性研究財団(FRB;www.fondationbiodiversite.fr)により支援されました。 J-CS は、この研究を VILLUM FONDEN(助成金 16549 号)の助成による VILLUM Investigator プロジェクト Biodiversity Dynamics in a Changing World への貢献と考えている。 7817>
利益相反声明
著者らは、本研究が利益相反の可能性があると解釈されるいかなる商業的または金銭的関係もない状態で行われたことを宣言している。 また、すべてのBIENデータ提供者(全リストはhttp://bien.nceas.ucsb.edu/bien/data-contributors/を参照)に感謝する。
補足資料
この論文の補足資料は、次のサイトで見ることができる。 https://www.frontiersin.org/articles/10.3389/fevo.2018.00219/full#supplementary-material
Banda, R.K., Delgado-Salinas, A., Dexter, K. G., Linares-Palomino, R., Oliveira-Filho, A., Prado, D., et al. (2016). 新熱帯乾燥林の植物多様性パターンとその保全の意味。 Science 353, 1383-1387. doi: 10.1126/science.aaf5080
PubMed Abstract | CrossRef Full Text | Google Scholar
Blonder, B., Lamanna, C., Violle, C., and Enquist, B. J. (2014). n次元のハイパーボリューム。 Glob. Ecol. Biogeogr. 23, 595-609. doi: 10.1111/geb.12146
CrossRef Full Text | Google Scholar
Blonder, B., Morrow, C. B., Maitner, B., Harris, D. J., Lamanna, C., Violle, C., et al(2018).。 n次元ハイパーボリュームを描画するための新しいアプローチ。 メソッズ・エコル。 Evol. 9, 305-319. doi: 10.1111/2041-210X.12865
CrossRef Full Text | Google Scholar
Bonan, G. B., Oleson, K. W., Fisher, R. A., Lasslop, G., and Reichstein, M. (2012). 葉の生理学的形質とキャノピーフラックスデータの調整:地域土地モデルバージョン4におけるTRYとFLUXNETデータベースの使用。 J. Geophys. Res. Biogeosci. 117:G02026. doi: 10.1029/2011JG001913
CrossRef Full Text | Google Scholar
Borregaard, M. K., and Rahbek, C. (2010). 地理的分布と種数量の関係の因果関係. Q. Rev. Biol. 85, 3-25. doi: 10.1086/650265
PubMed Abstract | CrossRef Full Text | Google Scholar
Bruggeman, J., Heringa, J., and Brandt, B. W. (2009). PhyloPars: Estimation of missing parameter values using phylogeny. Nucleic Acids Res. 37, W179-W184. doi: 10.1093/nar/gkp370
PubMed Abstract | CrossRef Full Text | Google Scholar
Cadotte, M. W., Carscadden, K., and Mirotchnick, N. (2011). 種を超えて:機能的多様性と生態学的プロセスおよびサービスの維持。 J. Appl. Ecol. 48, 1079-1087. doi: 10.1111/j.1365-2664.2011.02048.x
CrossRef Full Text | Google Scholar
Cornwell, W. K., Cornelissen, J. H., Amatangelo, K., Dorrepaal, E., Eviner, V. T., Godoy, O., and al. (2008). 植物種特性は、世界のバイオーム内のリター分解速度を支配している。 Ecol. Lett. 11, 1065-1071. doi: 10.1111/j.1461-0248.2008.01219.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Craine, J. M. (2009).植物種特性は、世界各地のバイオマス内のごみ分解速度を支配している。 野生植物の資源戦略. New Jersey, NJ: Princeton University Press. doi: 10.1515/9781400830640
CrossRef Full Text | Google Scholar
Currie, D. J., Mittelbach, G. G., Cornell, H. V., Field, R., Guégan, J. F., Hawkins, B. A., and al. (2004). 分類学的豊かさの広範な変動に関する気候に基づく仮説の予測と検証。 Ecol. Lett. 12, 1121-1134. doi: 10.1111/j.1461-0248.2004.00671.x
CrossRef Full Text | Google Scholar
de la Riva, E. G., Violle, C., Pérez-Ramos, I. M., Marañón, T., Navarro-Fernández, C. M., Olmo, M., et al(2018).のように。 多次元機能形質アプローチにより、地中海木質群集における環境ストレスの刷り込みが明らかになった。 Ecosystems 21, 248-262. doi: 10.1007/s10021-017-0147-7
CrossRef Full Text | Google Scholar
Díaz, S., Kattge, J., Cornelissen, J. H., Wright, I. J., Lavorel, S., Dray, S.、他 (2016).多次元的な機能特性アプローチにより、地中海の木質コミュニティの環境ストレスの影響が明らかになった。 植物の形態と機能のグローバルスペクトル。 Nature 529, 167-171. doi: 10.1038/nature16489
PubMed Abstract | CrossRef Full Text | Google Scholar
Díaz, S., Lavorel, S., de Bello, F., Quétier, F., Grigulis, K., and Robson, T.M. (2007). 生態系サービス評価における植物機能多様性の効果の取り込み。 Proc. Natl. Acad. Sci. U.S.A 104, 20684-20689. doi: 10.1073/pnas.0704716104
PubMed Abstract | CrossRef Full Text | Google Scholar
Donoghue, M. J. and Edwards, E. J. (2014). 植物におけるバイオームシフトとニッチ進化. Annu. Rev. Ecol. Evol.Syst. 45, 547-572. doi: 10.1146/annurev-ecolsys-120213-091905
CrossRef Full Text | Google Scholar
Douma, J. C., Aerts, R., Witte, J. P. M., Becker, R. M., Kunzmann, D., Metselaar, K.、他 (2012). 機能的に異なる植物形質の組み合わせは、北西ヨーロッパにおける広範な種群れを定量的に予測するための手段を提供する。 Ecography 35, 364-373. doi: 10.1111/j.1600-0587.2011.07068.x
CrossRef Full Text | Google Scholar
Engemann, K., Sandel, B., Enquist, B. J., Jørgensen, P. M., Kraft, N., Marcuse-Kubitza, A., et al.(2016).植物が持つ機能と形質の組み合わせは,ヨーロッパ北西部における広範な種群集を定量的に予測する手段となる. 西半球における植物機能群の支配のパターンとドライバー:膨大な植物学的データセットに基づくマクロ生態学的再評価。 Bot. J. Linn. Soc. 180, 141-160. doi: 10.1111/boj.12362
CrossRef Full Text | Google Scholar
Enquist, B. J., Condit, R., Peet, R., Thiers, B., and Schildhauer, M. (2016). The botanical information and ecology network (BIEN): Cyberinfrastructure for an integrated botanical information network to investigate the ecological impacts of global climate change on plant biodiversity.(植物情報生態ネットワーク). PeerJ Prepr. 4:e2615v2. doi: 10.7287/peerj.preprints.2615v2
CrossRef Full Text | Google Scholar
Garnier, E., Cortez, J., Billès, G., Navas, M. L., Roumet, C., Debussche, M., et al.(2004).を参照されたい。 植物機能マーカーが捉えた二次遷移時の生態系特性. Ecology 85, 2630-2637. doi: 10.1890/03-0799
CrossRef Full Text | Google Scholar
Goldsmith, G. R., Morueta-Holme, N., Sandel, B., Fitz, E. D., Fitz, S. D., Boyle, B., et al(2016).エコロジー85(1992).エコロジー85(1992).エコロジー85(1992).エコロジー85(1992).エコロジー86(1995).エコロジー86(1997)…. プラント・オーマティック アメリカ大陸の全植物へのダイナミックでモバイルなガイド。 メソッズ・エコル. Evol. 7, 960-965. doi: 10.1111/2041-210X.12548
CrossRef Full Text | Google Scholar
Goolsby, E. W., Bruggeman, J., and Ané, C. (2017). Rphylopars: missing data and within-species variationのための高速多変量系統比較法。 メソッズ・エコル. Evol. 8, 22-27. doi: 10.1111/2041-210X.12612
CrossRef Full Text | Google Scholar
Grenié, M., Denelle, P., Tucker, C. M., Munoz, F., and Violle, C. (2017). funrar: an R package to characterize functional rarity. Divers. Distrib. 23, 1365-1371. doi: 10.1111/ddi.12629
CrossRef Full Text | Google Scholar
Hortal, J., de Bello, F., Diniz-Filho, J. A. F., Lewinsohn, T. M., Lobo, J. M., and Ladle, R. J. (2015). 生物多様性の大規模な知識を悩ます7つの欠点。 Annu. Rev. Ecol. Evol. Syst. 46, 523-549. doi: 10.1146/annurev-ecolsys-112414-054400
CrossRef Full Text | Google Scholar
Kerkhoff, A.J., Enquist, B.J., Elser, J.J., and Fagan, W.F. (2005). 植物アロメトリー,化学量論と基礎生産性の温度依存性. Glob. Ecol. doi: 10.1111/j.1466-822X.2005.00187.x
CrossRef Full Text | Google Scholar
Kerkhoff, A. J., Fagan, W. F., Elser, J. J., and Enquist, B. J.(2006).,植物アロメトリー、ストイキオメトリー、温度依存性。 種子植物における窒素とリンのスケーリングにおける系統的および成長形態的変異. Am. Nat. 168, E103-E122. doi: 10.1086/507879
PubMed Abstract | CrossRef Full Text | Google Scholar
Kraft, N. J., Valencia, R., and Ackerly, D. D. (2008). アマゾンの森林における機能的形質とニッチに基づく樹木群集の形成。 Science 322, 580-582. doi: 10.1126/science.1160662
PubMed Abstract | CrossRef Full Text | Google Scholar
Lamanna, C., Blonder, B., Violle, C., Kraft, N. J., Sandel, B., et al.(2014).アマゾン林の機能形質と、ニッチに基づく樹木群集の形成。 機能的形質空間と緯度方向の多様性勾配. Proc. Natl.Acad. Sci. U.S.A. 111, 13745-13750. doi: 10.1073/pnas.1317722111
PubMed Abstract | CrossRef Full Text | Google Scholar
Lavorel, S., and Garnier, E. (2002). このような状況下において、植物形質から群集組成と生態系機能の変化を予測することは、聖杯の再検討につながる。 Funct. Ecol. 16, 545-556. doi: 10.1046/j.1365-2435.2002.00664.x
CrossRef Full Text | Google Scholar
Lavorel, S., Grigulis, K., Lamarque, P., Colace, M.-P., Garden, D., Girel, J.他 (2011).(英語). 複数の生態系サービスの景観分布を理解するための植物機能形質の利用。 J. Ecol. 99, 135-147. doi: 10.1111/j.1365-2745.2010.01753.x
CrossRef Full Text | Google Scholar
Li, Y., Shipley, B., Price, J. N., Dantas, V. de L., Tamme, R., Westoby, M., et al(2018).植物性サービス。 ハビタットフィルタリングが世界の植物群落の機能的ニッチ占有を決定している。 J. Ecol. 106, 1001-1009. doi: 10.1111/1365-2745.12802
CrossRef Full Text | Google Scholar
Magallón, S., Gómez-Acevedo, S., Sánchez-Reyes, L.L., and Hernández-Hernández, T. (2015). A metacalibrated time-tree documents the early rise of flowering plant phylogenetic diversity(メタキャリブレーションされたタイムツリーは顕花植物の系統的多様性の初期の上昇を記録している。 ニューフィトール. 207, 437-453. doi: 10.1111/nph.13264
PubMed Abstract | CrossRef Full Text | Google Scholar
Maitner, B. S., Boyle, B., Casler, N., Condit, R., Donoghue, J., Durán, S. M., et al (2018). The BIEN R package: a tool to access the Botanical information and ecology network (BIEN) database. Methods Ecol. Evol. 9, 373-379. doi: 10.1111/2041-210X.12861
CrossRef Full Text | Google Scholar
Moncrieff, G. R., Bond, W. J., and Higgins, S. I. (2016). 地球変動の影響を理解し予測するためのバイオーム概念の改訂. J. Biogeogr. 43, 863-873. doi: 10.1111/jbi.12701
CrossRef Full Text | Google Scholar
Novosolov, M., Rodda, G. H., North, A. C., Butchart, S. H. M., Tallowin, O. J. S., Gainsbury, A. M., et al. (2017). 個体群密度-範囲サイズ関係再考。 Glob. Ecol. Biogeogr. 26, 1088-1097. doi: 10.1111/geb.12617
CrossRef Full Text | Google Scholar
Olson, D. M., Dinerstein, E., Wikramanayake, E. D., Burgess, N. D., Powell, G. V. N., Underwood, E. C.、他 (2001). Terrestrial Ecoregions of the world: a new map of life on earth: a new global map of terrestrial ecoregions provides an innovative tool for conserving biodiversity. Bioscience 51, 933-938.doi: 10.1641/0006-3568(2001)0512.0.CO;2
CrossRef Full Text | Google Scholar
Pennington, R. T., Prado, D. E., and Pendry, C. A. (2000). 新熱帯の季節性乾燥林と第四紀の植生変化. J. Biogeogr. 27, 261-273. doi: 10.1046/j.1365-2699.2000.00397.x
CrossRef Full Text | Google Scholar
Pennington,R. T., Richardson,J. E.,and Lavin, M. (2006). このような状況下において,植物がどのように進化してきたのか,また,どのように進化してきたのか,さらに,どのように進化してきたのかを明らかにするために,植物系統樹,中立生態学,系統群構造から,種数の多いバイオームを歴史的に構築するための知見を得た。 ニューフィトール(New Phytol. 172, 605-616. doi: 10.1111/j.1469-8137.2006.01902.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Penone, C., Davidson, A. D., Shoemaker, K. T., Di Marco, M., Rondinini, C., Brooks, T. M., et al(2014).日本植物学会誌「植物と生物多様性」(2009)。 生命史的形質データセットにおける欠損データのインプット:どのアプローチが最もパフォーマンスが高いか? Methods Ecol. Evol. 5, 961-970. doi: 10.1111/2041-210X.12232
CrossRef Full Text | Google Scholar
Pillar, V. D., Blanco, C. C., Müller, S. C., Sosinski, E. E., Joner, F., and Duarte, L. D. S. (2013). 植物群落における機能的な冗長性と安定性. J. Veg. Sci. 24, 963-974. doi: 10.1111/jvs.12047
CrossRef Full Text | Google Scholar
Prado, D. E., and Gibbs, P. E. (1993). 南米の乾燥季節林における種分布のパターン. Ann. Mo. Bot. Gard. 80, 902-927. doi: 10.2307/2399937
CrossRef Full Text | Google Scholar
Qian, H., and Sandel, B. (2017)(2017). 北米の緯度・気候勾配を越えた地域的な被子植物群集の系統構造. Glob. Ecol. Biogeogr. 26, 1258-1269. doi: 10.1111/geb.12634
CrossRef Full Text | Google Scholar
Reich, P. B., Wright, I. J., Cavender-Bares, J., Craine, J. M., Oleksyn, J., Westoby, M., etc. (2003). 植物機能変異の進化:形質、スペクトル、戦略。 Int. J. Plant Sci. 164, S143-S164. doi: 10.1086/374368
CrossRef Full Text | Google Scholar
Silva de Miranda, P. L., Oliveira-Filho, A., Pennington, R. T., Neves, D. M., Baker, T. R., and Dexter, K.G. (2018).植物機能変異の進化とその特徴について Using tree species inventories to map biomes and assess their climatic overlaps in lowland tropical South America(南米低地のバイオームをマッピングし、その気候的重複を評価するための樹木種インベントリーの使用)。 Glob. Ecol. Biogeogr. 27, 899-912. doi: 10.1111/geb.12749
CrossRef Full Text | Google Scholar
Šímová, I., Violle, C., Kraft, N. J. B., Storch, D., Svenning, J.-C., Boyle, B.、他 (2015).KG. 北米の樹木群における形質平均と分散の変化:種の豊かさのパターンは機能空間と緩やかな関係にある。 Ecography 38, 649-658. doi: 10.1111/ecog.00867
CrossRef Full Text | Google Scholar
Šímová, I., Violle, C., Svenning, J.-C., Kattge, J., Engemann, K., Sandel, B., et al.(2018).樹木群における形質の平均と分散の変化:種の豊かさのパターンは機能空間と緩やかに関連している。 新世界における主要な植物形質の空間的パターンと気候の関係は、木本系と草本系で異なる。 J. Biogeogr. 45, 895-916. doi: 10.1111/jbi.13171
CrossRef Full Text | Google Scholar
Smith, S. A., and Beaulieu, J. M. (2009).新大陸の植物形質の空間パターンと気候との関係は、木本系と草本系で異なる。 生活史は顕花植物における気候ニッチ進化の速度に影響を与える。 Proc. Biol. Sci. 276, 4345-4352. doi: 10.1098/rspb.2009.1176
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, S.A., and Brown, J. W. (2018). 広く包括的な種子植物の系統樹を構築する。 Am. J. Bot. 105, 302-314. doi: 10.1002/ajb2.1019
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, S. A., and Donoghue, M. J. (2008). 分子進化の速度は顕花植物の生活史と連動している。 Science 322, 86-89. doi: 10.1126/science.1163197
PubMed Abstract | CrossRef Full Text | Google Scholar
Swenson, N.G. (2014). Phylogenetic imputation of plant functional trait databases.Ecography 37, 105-110. doi: 10.1111/j.1600-0587.2013.00528.x
CrossRef Full Text
Swenson, N. G., Enquist, B. J., Pither, J., Kerkhoff, A. J., Boyle, B., Weiser, M. D., et al. 南北アメリカにおける木本植物機能多様性の生物地理学とフィルタリング. Glob. Ecol. Biogeogr. 21, 798-808. doi: 10.1111/j.1466-8238.2011.00727.x
CrossRef Full Text | Google Scholar
Swenson, N. G., Weiser, M. D., Mao, L., Araújo, M. B. , Diniz-Filho, J. A. F., Kollmann, J. , et al. (2017). 新規大陸設定にわたる系統と樹木機能多様性の予測。 Glob. Ecol. Biogeogr. 26, 553-562. doi: 10.1111/geb.12559
CrossRef Full Text | Google Scholar
Umaña, M. N., Mi, X., Cao, M., Enquist, B. J., Hao, Z., Howe, R.、その他 (2017).P. (2017)。 樹木の希少性の説明における機能的独自性と空間的集約の役割。 Glob. Ecol. Biogeogr. 26, 777-786. doi: 10.1111/geb.12583
CrossRef Full Text | Google Scholar
van Bodegom, P. M., Douma, J. C., and Verheijen, L. M. (2014). 地球規模の植生分布をモデル化するための完全形質ベースのアプローチ。 Proc. Natl.Acad. Sci. U.S.A. 111, 13733-13738. doi: 10.1073/pnas.1304551110
PubMed Abstract | CrossRef Full Text | Google Scholar
Violle, C., Reich, P. B., Pacala, S.W., Enquist, B.J., and Kattge, J.(2014年). 機能的生物地理学の出現と将来性. Proc. Natl.Acad. Sci. U.S.A. 111, 13690-13696. doi: 10.1073/pnas.1415442111
PubMed Abstract | CrossRef Full Text | Google Scholar
Violle, C., Thuiller, W., Mouquet, N., Munoz, F., Kraft, N. J. B., Cadotte, M. W., et al. (2017). 機能的希少性:外れ値の生態学。 Trends Ecol. Evol. 32, 356-367. doi: 10.1016/j.tree.2017.02.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Walker, B., Kinzig, A., and Langridge, J. (1999). 植物属性の多様性、回復力、および生態系機能:優占種と少数種の性質と意義。 Ecosystems 2, 95-113. doi: 10.1007/s100219900062
CrossRef Full Text | Google Scholar
Westoby, M. (1998). リーフハイトシード(LHS)植物生態学的戦略スキーム。 地球気候と植物バイオームの分布. Philos. Trans. R. Soc. Lond. B. Biol. 359, 1465-1476. doi: 10.1098/rstb.2004.1525
PubMed Abstract | Cross Ref Full Text | Google Scholar
Wright, I. J., Reich, P. B., Westoby, M., Ackerly, D. D., Baruch, Z., Bongers, F., et al. 世界的な葉の経済スペクトル。 Nature 428, 821-827. doi: 10.1038/nature02403
PubMed Abstract | CrossRef Full Text | Google Scholar
Xia, J., Niu, S., Ciais, P., Janssens, I. A., Chen, J., Ammann, C., and al. (2015). 植物のフェノロジーと生理による陸域総一次生産性の共同制御. Proc. Natl.Acad. Sci. U.S.A. 112, 2788-2793. doi: 10.1073/pnas.1413090112
PubMed Abstract | CrossRef Full Text | Google Scholar
Zanne, A.E., Pearse, W.D., Cornwell, W.K., McGlinn, D.J., Wright, I.J., and Uyeda, J.C. (2018).のような、植物が持つフェノロジーと生理学による陸域の総定量生産性の共同制御。 被子植物の機能的生物地理学:極限での生活. New Phytol. 218, 1697-1709. doi: 10.1111/nph.15114
PubMed Abstract | CrossRef Full Text | Google Scholar
Zanne, A. E., Tank, D. C., Cornwell, W. K., Eastman, J. M., Smith, S. A., FitzJohn, R. G., and al. (2014). 被子植物の凍結環境への放射に関する3つの鍵。 Nature 506, 89-92. doi: 10.1038/nature12872
PubMed Abstract | CrossRef Full Text | Google Scholar
.