- Introducere
- Metode
- Date de distribuție și clasificare a biomului
- Compoziția speciilor între biome
- Date privind trăsăturile
- Măsurarea hipervolumelor trăsăturilor
- Distinctivitatea funcțională și răspândirea
- Spațiu funcțional și similaritate între biome
- Rezultate
- Discuție
- Cunoștințe insuficiente
- Planuri comune în cadrul biomurilor
- Comparații între biomi
- Concluzii
- Date disponibile
- Contribuții ale autorilor
- Finanțare
- Declarație privind conflictul de interese
- Recunoștințe
- Supplementary Material
Introducere
Ecologii și biogeografii organizează biodiversitatea plantelor terestre în biomuri determinate de climă, cu fizionomii caracterizate de formele de creștere și trăsăturile funcționale ale speciilor dominante (Moncrieff et al., 2016). Într-adevăr, conceptul de biom are o istorie îndelungată care datează de la Carl Ludwig Willdenow și Alexander von Humboldt. Willdenow a recunoscut că climatele similare susțin forme de vegetație similare, iar Humboldt a observat asocierea generalizată dintre distribuția plantelor, fizionomia și factorii de mediu. În general, conceptul de biom reflectă ipoteza că presiunile de mediu similare selectează specii cu atribute funcționale similare, independent de istoria lor evolutivă. În același timp, biomii existenți ai Pământului sunt, într-o oarecare măsură, distincți din punct de vedere filogenetic, multe/majoritatea speciilor caracteristice provenind din anumite linii genealogice care prezintă adaptări esențiale nu doar la selecția climatică, ci și la presiuni suplimentare, cum ar fi focul sau megaherbivorele (Woodward et al., 2004; Pennington et al., 2006; Donoghue și Edwards, 2014). Deoarece biomii reprezintă regularități la scară largă în vegetația Pământului, înțelegerea diferențelor funcționale dintre biomi este extrem de importantă pentru modelarea ciclului global al carbonului și a funcționării sistemului Pământului, inclusiv a răspunsurilor la schimbările globale antropice (Bonan et al, 2012; van Bodegom et al., 2014; Xia et al., 2015).
În ultimele două decenii s-a înregistrat o creștere rapidă a înțelegerii dimensiunii funcționale a biodiversității vegetale la scară continentală (Swenson et al., 2012; Lamanna et al., 2014; Šímová et al., 2018). Eforturile extinse de sinteză și analiză a datelor au definit un spațiu limitat de trăsături funcționale care încorporează variația în dimensiunea generală a plantelor, economia frunzelor și hidraulica, cunoscute sub numele de spectre de trăsături funcționale (Wright et al., 2004; Díaz et al., 2016). Spectrele trăsăturilor funcționale definesc compromisurile fiziologice și ecologice care determină strategiile de istorie a vieții plantelor (Westoby, 1998; Reich et al., 2003; Craine, 2009) și influența acestora asupra ansamblului comunității (Kraft et al., 2008), a funcției ecosistemului (Lavorel și Garnier, 2002; Garnier et al., 2004; Kerkhoff et al., 2005; Cornwell et al., 2008) și chiar a ratelor de evoluție (Smith și Donoghue, 2008; Smith și Beaulieu, 2009). Generalitatea și utilitatea spectrelor de trăsături funcționale ale plantelor au grăbit încorporarea lor în modelele de distribuție globală a vegetației (van Bodegom et al., 2014), în ciclurile biogeochimice (Bonan et al., 2012) și în serviciile ecosistemice (Díaz et al., 2007; Cadotte et al., 2011; Lavorel et al., 2011). Cu toate acestea, puține studii au examinat măsuri cuprinzătoare ale diversității funcționale la scara biomului, atât pentru speciile și formele de viață dominante, cât și pentru cele subordonate.
Există două seturi contrastante de predicții cu privire la modul în care diversitatea funcțională variază între biomele distincte din punct de vedere climatic și fizionomic. Pe de o parte, dacă biodiversitatea biomurilor reflectă variația spațiului de nișă ecologică disponibil, biomurile mai puțin diverse din punct de vedere taxonomic ar trebui să reprezinte un subansamblu mai mic, în mare parte imbricate, al spațiului funcțional ocupat de biomurile mai diverse. Pe de altă parte, datorită naturii globale a compromisurilor fundamentale în structura și funcția plantelor și a presiunilor de selecție similare care acționează în mod similar între specii, ansamblurile care ocupă medii diferite pot, de fapt, să împartă zone similare ale spațiului trăsăturilor (Reich et al., 2003; Wright et al., 2004; Díaz et al., 2016). Studii recente confirmă faptul că relațiile dintre climă și diversitatea funcțională și taxonomică (de exemplu, bogăția de specii) sunt complexe și dependente de scară. În unele cazuri, diversitatea funcțională urmărește îndeaproape gradienții climatici, cu o diversitate funcțională mai scăzută în condiții mai variabile și extreme (Swenson et al., 2012; de la Riva et al., 2018), iar volumul spațiului funcțional în ansamblurile locale se extinde odată cu creșterea bogăției taxonomice (Lamanna et al., 2014; Li et al., 2018). Cu toate acestea, analizele bazinelor de specii la scară regională sugerează că schimbările mari în bogăția de specii pot fi asociate cu un impact minim asupra diversității funcționale (Lamanna et al., 2014; Šímová et al., 2015). Mai mult, răspunsurile diversității taxonomice și funcționale la climă pot diferi între principalele forme de creștere a plantelor (Šímová et al., 2018), iar diversitatea relativă a diferitelor forme de creștere se poate schimba substanțial de-a lungul gradienților climatici care definesc diferite biome (Engemann et al., 2016).
Progresul în analiza diversității funcționale la scară continentală până la scară globală a fost limitat în parte de deficitul de date în catalogarea aspectelor taxonomice, distribuționale, filogenetice și funcționale ale biodiversității (Hortal et al., 2015). Mai mult, chiar și având în vedere datele limitate disponibile, există provocări informatice semnificative asociate cu standardizarea și integrarea unor seturi de date mari și disparate care descriu distribuțiile geografice, trăsăturile funcționale și relațiile filogenetice ale speciilor (Violle et al., 2014). Aici examinăm distribuția diversității funcționale a speciilor de plante în biomurile din America de Nord și America de Sud, utilizând baza de date Botanical Information and Ecology Network (BIEN; Enquist et al., 2016; Maitner et al., 2018), care reunește informații privind distribuția și trăsăturile funcționale pentru >100.000 de specii de plante terestre. În mod specific, examinăm variația diversității funcționale și a caracterului distinctiv al biomurilor, pe baza hărților de distribuție a speciilor de plante terestre la o scară de celule de grilă de 100 × 100 km și a unui set de date cuprinzător de șase trăsături funcționale care reflectă axele majore de variație a strategiilor ecologice.
Obiectivele noastre în acest studiu sunt de trei ori. În primul rând, pentru a evidenția deficiențele persistente ale datelor, documentăm amploarea datelor disponibile care caracterizează diversitatea funcțională și caracterul distinctiv al biomurilor. În al doilea rând, având în vedere datele disponibile, caracterizăm distribuția diversității funcționale în cadrul biomurilor, atât pentru formele de creștere dominante, cât și pentru cele subordonate. Aceste analize ne permit să cuantificăm mai bine distinctivitatea funcțională a unui biom prin identificarea celor mai comune strategii funcționale ale celor mai răspândite specii din cadrul acestuia. În al treilea rând, ne întrebăm dacă biomii sunt de fapt caracterizați de colecții de specii distincte din punct de vedere funcțional, pe baza suprapunerii hipervolumelor multidimensionale în spațiul trăsăturilor funcționale.
Metode
Date de distribuție și clasificare a biomului
Pentru a reduce efectele prejudecăților de eșantionare caracteristice seturilor de date de prezență compilate din mai multe resurse, am folosit hărțile de distribuție BIEN 2.0 pentru 88.417 de specii de plante distribuite în America de Nord și de Sud (Goldsmith et al., 2016). Baza de date BIEN integrează observații standardizate ale plantelor provenite din specimene de ierbar și din inventare ale parcelelor de vegetație. Hărțile de distribuție a speciilor din bazele de date BIEN au fost estimate folosind una dintre cele trei abordări, în funcție de prezențele disponibile pentru fiecare specie. Pentru speciile cu doar una sau două prezențe (c. 35 % din specii), aria de răspândire geografică a fost definită ca o zonă pătrată de 75.000 km2 în jurul fiecărui punct de date. Pentru speciile cu trei sau patru prezențe, ariile geografice au fost identificate cu ajutorul unui hull convex (c. 15% din specii). În cele din urmă, hărțile ariei de răspândire pentru speciile cu cel puțin cinci prezențe au fost obținute cu ajutorul algoritmului de modelare a distribuției speciilor Maxent, folosind 19 straturi climatice ca variabile predictive și 19 vectori proprii spațiali ca filtre pentru a constrânge previziunile excesive ale modelelor (a se vedea Goldsmith et al, 2016 pentru mai multe detalii privind metodologia hărților de distribuție).
Am suprapus hărțile de specii de plante BIEN 2.0 pe o hartă grilă de 100 × 100 km cu o proiecție Lambert Azimuthal Equal Area pentru a obține o matrice de prezență/absență a speciilor pentru fiecare celulă de grilă. Pe baza clasificării biomului Olson et al. (2001), am atribuit fiecare celulă de grilă a matricei la una dintre categoriile de biom. Din cauza limitărilor de calcul pentru analizele ulterioare, am alăturat unele biome pe baza similitudinilor lor în ceea ce privește clima și vegetația și literatura de specialitate pentru a obține o clasificare largă, așa cum este descrisă în tabelul 1 (cu excepția apelor interioare, a rocilor și gheții și a mangrovelor). Ecoregiunile Chaco și Caatinga au fost clasificate ca păduri xerice și, respectiv, păduri uscate, urmând Prado și Gibbs (1993), Pennington et al. (2000), Banda et al. (2016), Silva de Miranda et al. (2018).
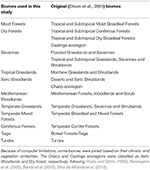
Tabel 1. Prezentare generală a clasificării biomului adoptat în acest studiu și a clasificării echivalente (Olson et al., 2001) a biomurilor (cu excepția apelor interioare, a rocilor și gheții și a mangrovelor).
Compoziția speciilor între biome
>Pentru a înțelege variația în spațiul trăsăturilor funcționale între biome, am explorat mai întâi diferențele în compoziția speciilor de plante între ele. Pe baza listei de specii pentru fiecare celulă de grilă și biom, am definit speciile caracteristice fiecărui biom ca fiind cele care au proporția maximă a arealului lor în biomul respectiv. Aceste liste de specii predominante din punct de vedere geografic într-un anumit biom au fost comparate cu lista de specii din alte biome. Această comparație pe perechi oferă o modalitate simplă de a deduce o suprapunere taxonomică direcțională între biome (și anume, proporția de specii predominante dintr-un biom împărtășite cu un alt biom). Din cele 88.417 specii cu hărți ale arealului disponibile, 44.899 de specii au areale care acoperă mai mult de un biom, iar 43.518 specii sunt endemice pentru un anumit biom.
Date privind trăsăturile
Am extras toate informațiile privind trăsăturile pentru speciile de plante din Lumea Nouă disponibile în setul de date BIEN 3.0 (recuperate la 7 februarie 2018), ceea ce dă un total de 80.405 observații privind trăsăturile la nivelul speciilor. Apoi am filtrat informațiile pentru șase trăsături funcționale: înălțimea maximă a plantelor (m), masa semințelor (mg), densitatea lemnului (mg/cm3), suprafața specifică a frunzelor SLA (cm2/g) și concentrația de fosfor din frunze și de azot din frunze pe unitate de masă (mg/g). În total, după filtrare au rămas 18 192 de observații la nivel de specie. Dintre acestea, subsetul de 8.820 de specii cu hărți ale arealului și informații despre trăsături a fost utilizat pentru analize ulterioare.
Pentru a estima spațiul trăsăturilor funcționale pentru fiecare biom, am avut nevoie de date complete despre trăsături. Din acest motiv, am imputat filogenetic datele de trăsături lipsă (Bruggeman et al., 2009; Penone et al., 2014; Swenson, 2014; Swenson et al., 2017) folosind pachetul R “Rphylopars” v 0.2.9 (Goolsby et al., 2017) și filogenia recent publicată a plantelor cu semințe de către Smith și Brown (2018) ca bază de referință. Imputarea filogenetică este un instrument de predicție a datelor lipsă în seturile de date privind trăsăturile funcționale, bazat pe ipoteza că speciile strâns înrudite tind să aibă valori similare ale trăsăturilor (Swenson, 2014). Am utilizat arborele ALLBM (adică banca de gene și taxonul Open Tree of life cu o coloană vertebrală furnizată de Magallón et al., 2015) deoarece a maximizat suprapunerea între speciile cu informații disponibile despre trăsături și distribuție. După imputarea trăsăturilor, un total de 7.842 de specii cu informații complete despre trăsături și hărți de distribuție au rămas pentru analize ulterioare.
Măsurarea hipervolumelor trăsăturilor
Am măsurat diversitatea funcțională relativă a biomurilor prin calcularea hipervolumelor trăsăturilor din bazinele de specii din cadrul celulelor biomurilor. Din cauza limitărilor computaționale, am construit hipervolumele trăsăturilor folosind un eșantion aleatoriu de 20% din celulele fiecărui biom. Hipervolumele pentru fiecare celulă de grilă au fost estimate folosind cele șase trăsături funcționale extrase și pachetul R “hypervolume” (Blonder et al., 2014, 2018), utilizând metoda Gaussian KDE cu estimatorul implicit de lățime de bandă Silverman. Masa semințelor, înălțimea și densitatea lemnului au fost transformate în logaritmi, în timp ce SLA a fost transformată cu rădăcină pătrată. Toate trăsăturile au fost scalate și centrate înainte de analiză. Hipervolumele sunt raportate în unități de deviații standard (sd) la puterea la numărul de trăsături utilizate (adică, sd6).
Distinctivitatea funcțională și răspândirea
Pentru că biomii sunt caracterizați de vegetația lor dominantă, am examinat, de asemenea, răspândirea geografică și distinctivitatea funcțională a speciilor din cadrul biomilor. Utilizând hărțile de răspândire a speciilor și informațiile privind trăsăturile funcționale, am estimat distinctivitatea funcțională și răspândirea fiecărei specii în fiecare biom, urmând cadrul conceptual al rarității funcționale de către Violle et al. (2017). Utilizând întregul set de trăsături (măsurate și imputate), am măsurat mai întâi distanța euclidiană în spațiul trăsăturilor standardizate între toate perechile de specii. Apoi, am calculat distinctivitatea funcțională (Di) pentru fiecare specie din fiecare biom ca fiind distanța funcțională medie a unei specii față de N alte specii din fondul de specii din biom. Di a fost scalat între 0 și 1 în cadrul fiecărui biom, valorile mai mici reprezentând speciile care sunt comune din punct de vedere funcțional (redundante) și valorile mai mari reprezentând speciile care sunt distincte din punct de vedere funcțional, în comparație cu celelalte specii din biom. De asemenea, am estimat răspândirea geografică (Wi) a unei specii într-un biom, măsurată ca număr de celule de grilă ocupate de specia respectivă în cadrul unui biom, raportat la numărul total de celule din biomul respectiv. O valoare de 1 indică faptul că specia focală este prezentă în toate celulele din grilă acoperite de biom. Atât distinctivitatea funcțională, cât și răspândirea geografică au fost calculate cu ajutorul pachetului R “funrar” (Grenié et al., 2017).
Pentru că nu dispunem de măsuri cuprinzătoare de dominanță bazate pe abundența sau biomasa locală, am utilizat măsurile Di și Wi pentru a identifica pentru fiecare biom cele mai “comune” specii ca fiind cele care sunt atât răspândite geografic (Wi > 0,5), cât și similare din punct de vedere funcțional (Di < 0,25). Acest ultim prag a fost utilizat deoarece cel de-al treilea cuantile al valorilor distinctivității funcționale între biome a variat de la 0,2 la 0,3. Pentru a discrimina speciile distincte din punct de vedere funcțional față de speciile similare din punct de vedere funcțional în analizele ulterioare, am folosit, prin urmare, o valoare de 0,25 ca prag (adică, speciile cu valori ale distinctivității < 0,25 au fost considerate comune sau redundante din punct de vedere funcțional în cadrul unui anumit biom).
Spațiu funcțional și similaritate între biome
Pentru a estima suprapunerea între hipervolumele biomurilor am utilizat indicele de similaritate Sørensen folosind (i) numărul total de specii și (ii) lista de specii considerate comune din punct de vedere funcțional și răspândite geografic pentru fiecare biom. Similitudinea spațială a trăsăturilor funcționale a fost calculată ca fiind suprapunerea fracționară pe perechi a hipervolumelor dintre biomi. Suprapunerea fracționară a fost calculată prin împărțirea de două ori a volumului intersecției a două hipervolumuri la volumul uniunilor acestora. Toate hipervolumele au fost estimate cu ajutorul pachetului R “hypervolume” (Blonder et al., 2014, 2018), folosind metoda Gaussian KDE cu estimatorul implicit al lățimii de bandă Silverman.
Rezultate
Am constatat o suprapunere substanțială în compoziția speciilor de plante între biomi, pe baza intersecției intervalelor de specii modelate (a se vedea figura 1 și tabelul suplimentar 1). Această suprapunere taxonomică este mai mare în cadrul biomurilor tropicale și temperate, relativ puține specii fiind partajate între aceste două zone climatice. Este interesant faptul că pădurile xerofile împart specii atât cu biomul tropical, cât și cu cel temperat. În ciuda proporției ridicate de specii caracteristice pădurilor tropicale umede (83 %), acest biom împarte, de asemenea, un număr mare de specii cu alte biome tropicale, cum ar fi pădurile uscate, savanele, pădurile xerice și pajiștile tropicale. Suprapunerea taxonomică ridicată a biomurilor temperate, cum ar fi pajiștile temperate, pădurile mediteraneene, taiga și tundra, sugerează potențialul unei distinctivități funcționale scăzute, unii biomi temperați reprezentând un subansamblu mai slab al biomurilor tropicale mai bogate în specii și mai diverse din punct de vedere funcțional.
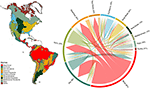
Figura 1. Suprapunerea speciilor de plante între biomii din Lumea Nouă. Valorile procentuale exprimă fracțiunea de specii care apar într-un biom și care au cea mai mare proporție a arealului lor geografic în acel biom, N = 84.413. Baza fiecărei săgeți ramificate este poziționată astfel încât să arate biomul care include cea mai mare proporție din aria de răspândire a unei specii, în timp ce lățimea reprezintă numărul de specii împărtășite cu biomurile aflate la vârfurile săgeții. A se vedea Tabelul suplimentar 1 pentru matricea subiacentă.
Cu mai puțin de 10% din speciile cartografiate reprezentate, datele privind trăsăturile au fost destul de rare pentru ansamblurile noastre de specii la scară biologică (Figura 2A). Mai mult, datele disponibile privind trăsăturile au variat substanțial între trăsături, clade de plante și biome. Din punct de vedere filogenetic, P-ul frunzelor și densitatea lemnului au fost deosebit de slab reprezentate, clade întregi fiind lipsite de date disponibile. Prin urmare, din cele aproximativ 2/3 din valorile trăsăturilor pentru cele 7 842 de specii ale noastre care au trebuit să fie imputate, multe au trebuit să fie atribuite în absența unor taxoni strâns înrudiți. Din punct de vedere geografic, SLA, înălțimea și masa semințelor sunt subeșantionate la tropice și în zona temperată a Americii de Sud, în timp ce N din frunze și densitatea lemnului sunt eșantionate mai uniform între regiuni. P din frunze este slab eșantionat în toate biomele. Hipervolumele de trăsături create din bazinele de specii ale unei selecții aleatorii de 20 % din celule pentru fiecare biom prezintă în principal diferențe între biomul tropical și cel temperat/ rece (Figura 2B). Hipervolumele trăsăturilor tind să fie mai mari și mai variabile în biomurile tropicale, cu cea mai mare variație în pădurile tropicale umede și uscate. În rândul biomurilor temperate/ reci, pădurile de conifere și pădurile mixte temperate susțin cele mai mari hipervolumuri de trăsături. În mod interesant, tufișurile xerice prezintă o distribuție a hipervolumelor mai asemănătoare cu cea a biomurilor tropicale decât cu cea a biomurilor temperate.
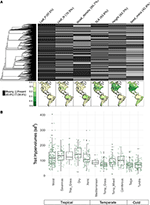
Figura 2. (A) Proporția de specii din baza de date BIEN 3.0 cu valori ale trăsăturilor cunoscute (gri) sau lipsă (negru). Filogenia din stânga corespunde arborelui ALLBM al plantelor de semințe de la Smith și Brown (2018) (de exemplu, GenBank și Open Tree of life taxa cu o coloană vertebrală furnizată de Magallón et al., 2015). Hărțile reprezintă proporția de specii cu informații despre trăsături în raport cu numărul total de specii din baza de date BIEN 2.0. (B) Distribuția hipervolumelor de trăsături din 20 % din celulele de 100 × 100 km selectate aleatoriu în fiecare biom. Hipervolumele sunt raportate în unități de deviații standard la puterea numărului de trăsături utilizate.
Relația dintre distinctivitatea funcțională și distribuția geografică este remarcabil de similară între biomi (Figura 3). În fiecare biom, marea majoritate a speciilor sunt atât restrânse din punct de vedere geografic, cât și similare din punct de vedere funcțional. În același timp, speciile cele mai distincte din punct de vedere funcțional din cadrul fiecărui biom au fost, în general, de asemenea, restrânse din punct de vedere geografic, iar speciile cele mai răspândite din punct de vedere geografic au fost aproape întotdeauna similare din punct de vedere funcțional.
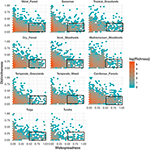
Figura 3. Modele de distinctivitate funcțională între biomi. Distinctivitatea reprezintă modul în care speciile sunt distanțate între ele din punct de vedere funcțional în cadrul unui biom (de exemplu, distanța fenotipică medie pe perechi dintre o specie focală și toate celelalte). Cu cât valoarea este mai mare, cu atât o specie este mai îndepărtată de centroidul spațiului funcțional al biomului. Răspândirea măsoară cât de comună este o specie din punct de vedere geografic în cadrul unui biom. O valoare de 0 indică faptul că o specie este prezentă într-o singură celulă a biomului. Valorile Bin reprezintă numărul de specii pe o scară logică. Dreptunghiurile reprezintă speciile care sunt considerate similare din punct de vedere funcțional și răspândite în fiecare biom, folosind valori limită de 0,25 și 0,5 ale distinctivității și, respectiv, ale răspândirii.
Am constatat o variație considerabilă în distribuția proporțională a formelor de creștere în cadrul biomului și între biomi. Speciile lemnoase reprezintă formele de creștere dominante în biomurile tropicale în ceea ce privește numărul de specii, în timp ce speciile erbacee domină în mediile temperate (figura 4). Atunci când am luat în considerare doar acele specii care sunt răspândite și comune din punct de vedere funcțional în fiecare biom, distribuția formelor de creștere între biomi s-a schimbat, în special în ceea ce privește proporția de arbori, ierburi și ierburi. De exemplu, în biomurile tropicale, proporția de arbori a scăzut în fiecare biom, cu excepția pădurilor umede. În biomurile temperate, proporția de ierburi a crescut, în special spre regiunile polare. Este interesant faptul că distribuția formelor de creștere în pădurile xerice se aseamănă mai mult cu distribuția din biomurile temperate atunci când sunt luate în considerare doar speciile răspândite și comune din punct de vedere funcțional.
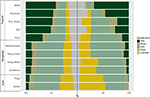
Figura 4. Distribuția formelor de creștere în fiecare biom (stânga) utilizând numărul total de specii și (dreapta) utilizând numai acele specii care sunt similare din punct de vedere funcțional și răspândite în cadrul biomului (a se vedea figura 3).
Comparațiile pe perechi ale compoziției speciilor între biomi relevă trei grupuri principale care reprezintă zonele climatice tropicale, temperate și polare (figura 5A), reflectând suprapunerea taxonomică ridicată în cadrul acestor regiuni, dar nu și între acestea (figura 1). Comparațiile pe perechi ale hipervolumelor trăsăturilor între biome arată o grupare mai puțin clară a zonelor climatice (Figura 5B). În acest caz, spațiul funcțional al pădurilor xerofile se suprapune în mod semnificativ cu biomurile temperate. Această analiză relevă, de asemenea, suprapunerea spațiului funcțional al taigalei cu pajiștile temperate și cu pădurile mixte. Comparația pe perechi a hipervolumelor de trăsături între biomi, utilizând doar acele specii considerate comune și răspândite din punct de vedere funcțional, arată o suprapunere mai mică a spațiilor de trăsături între și în cadrul zonelor climatice (figura 5C). Cu toate acestea, chiar dacă pădurile xerice sunt acum grupate cu restul biomilor tropicali, aceste habitate, împreună cu pajiștile tropicale, prezintă o mare suprapunere în spațiul funcțional cu biomii temperați, cum ar fi pădurile mediteraneene și pajiștile temperate.
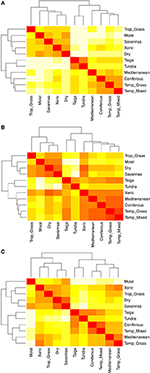
Figura 5. (A) Disimilaritatea perechilor în ceea ce privește compoziția speciilor între biome. (B) Disimilaritatea pe perechi în hipervolumele trăsăturilor (similaritate 1-Sørensen) între biomi folosind numărul total de specii. (C) Disimilaritatea perechilor în hipervolumele trăsăturilor (similaritate 1-Sørensen) între biomi, utilizând numai acele specii care sunt considerate similare din punct de vedere funcțional și răspândite. Cu cât celula este mai deschisă, cu atât disimilaritatea este mai mare.
Discuție
Analizele noastre oferă trei perspective importante pentru înțelegerea biomurilor terestre printr-o lentilă funcțională. În primul rând, arătăm că, în ciuda progreselor înregistrate în compilarea și sinteza datelor primare privind biodiversitatea, persistă lacune semnificative în materie de cunoștințe care pot limita capacitatea noastră de a cuantifica biodiversitatea funcțională a biomurilor la scară continentală până la scară globală. În al doilea rând, analizele noastre asupra datelor disponibile arată totuși că toți biomii din America de Nord și de Sud prezintă un model comun remarcabil, în care speciile cele mai răspândite din punct de vedere geografic din orice biom tind să împărtășească trăsături funcționale comune, în timp ce speciile cele mai distincte din punct de vedere funcțional sunt invariabil restrânse în distribuția lor. În al treilea rând, atunci când sunt luate în considerare doar speciile răspândite și comune din punct de vedere funcțional, biomurile pot fi mai ușor de distins din punct de vedere funcțional, iar modelele de disimilaritate între biomuri par să reflecte o corespondență între climă și spațiul de nișă funcțional al plantelor. Luate împreună, rezultatele noastre sugerează că, deși studiul diversității funcționale a biomurilor se află încă în stadii de formare, dezvoltarea în continuare a acestui domeniu va oferi probabil informații care să facă legătura între evoluție, biogeografie, asamblarea comunităților și funcția ecosistemică.
Cunoștințe insuficiente
Baza de date BIEN este special concepută pentru a acoperi lacunele din cunoștințele noastre despre biodiversitatea vegetală, însă, după cum subliniază Hortal et al. (2015), interacțiunea lacunelor de cunoștințe duce la incertitudine în cuantificarea biodiversității la cele mai mari scări. De exemplu, nu numai că mai puțin de 10 % din speciile noastre cartografiate aveau disponibile atât date privind trăsăturile, cât și informații filogenetice, dar acele date lipsă privind trăsăturile (~66 % din valorile trăsăturilor speciilor) erau destul de structurate, atât din punct de vedere filogenetic, cât și geografic. Astfel, deficitul raunkiaerian în ceea ce privește datele privind trăsăturile funcționale (sensu Hortal et al., 2015) interacționează cu deficitul wallacean în ceea ce privește informațiile privind distribuția și cu deficitul darwinist în ceea ce privește înțelegerea filogenetică. Și, deși putem utiliza cunoștințele filogenetice pentru a imputa în mod rezonabil aceste valori lipsă (Swenson et al., 2017), mai multe aspecte ale modelelor rezultate care sunt importante pentru clasificarea biomului rămân incerte. De exemplu, datele existente privind formele de creștere sugerează că speciile lemnoase domină în toate biomurile tropicale, în timp ce diversitatea proporțională a formelor de creștere erbacee este mult mai mare în biomurile temperate și polare (a se vedea, de asemenea, Engemann et al., 2016; Zanne et al., 2018). Această creștere latitudinală a diversității relative a plantelor erbacee reflectă prejudecățile de eșantionare și lipsa de cunoștințe taxonomice despre plantele erbacee tropicale sau reflectă istoriile evolutive și biogeografice diferite din regnurile nearctic și neotropical? Formele de creștere dominante sunt una dintre principalele caracteristici distinctive ale biomurilor, astfel încât schimbările sistematice în cele mai diverse forme de creștere (fie că sunt dominante sau nu) vor influența în mod necesar previziunile noastre cu privire la modul în care diversitatea funcțională s-ar putea schimba în biomi și la modul în care aceste schimbări vor afecta funcția și serviciile ecosistemice.
Am folosit hărțile de distribuție a speciilor ca date de intrare, în principal pentru a depăși problema incontestabilă a prejudecăților de eșantionare care este caracteristică seturilor de date compilate din mai multe surse (a se vedea figura suplimentară 1). Cu toate acestea, hărțile de areal întocmite din modele de distribuție a speciilor ar putea reprezenta un element suplimentar de incertitudine în rezultatele noastre. De exemplu, având în vedere rezoluția geografică a acestui studiu și complexitatea spațială a anumitor biome, aceste modele ar fi putut supraestima extinderea geografică a unor specii de la celulele unui singur biom la celule ale unor biome apropiate și interdigitate. Utilizarea datelor de prezență nu a schimbat rezultatele generale ale acestui studiu (a se vedea figurile suplimentare 2-6); cu toate acestea, o ușoară scădere a suprapunerii funcționale între biome ar putea într-adevăr să dezvăluie o supraestimare a ariei de răspândire a unor specii (a se vedea figurile suplimentare 2, 5 și 6). Utilizarea informațiilor privind prezența a modificat, de asemenea, relațiile generale dintre distinctivitatea funcțională și răspândirea în rezultatele noastre, din cauza supraeșantionării relative a regiunilor temperate față de cele tropicale (a se vedea figura suplimentară 4). În acest caz, măsura noastră de răspândire este limitată de eșantionarea extrem de limitată a majorității speciilor din biomul tropical. Pe măsură ce seturile de date și eșantionarea se îmbunătățesc, este posibil să existe posibilități de a reduce aceste incertitudini în viitor.
O altă lacună de date care trebuie abordată pentru a înțelege diversitatea funcțională a biomurilor este deficitul prestonian de informații privind abundența. Abundența este deosebit de importantă în studiile privind diversitatea funcțională, deoarece trăsăturile care contează cel mai mult pentru asamblarea comunității, funcționarea ecosistemului și ciclul biogeochimic sunt cele ale celor mai dominante specii. Deoarece ne-am bazat ansamblurile noastre pe hărțile de distribuție derivate din modelele de distribuție a speciilor, am putut aborda doar componenta geografică a răspândirii, dar relația pozitivă dintre abundența locală și distribuția geografică, așa-numita relație de ocupare-abundență, sugerează că speciile cele mai răspândite în fiecare biom pot fi frecvent și printre cele mai abundente (Borregaard și Rahbek, 2010; Novosolov et al., 2017). Deoarece biomii sunt, în general, caracterizați de formele de creștere dominante în regiune, integrarea informațiilor privind abundența ar putea avea ca rezultat biomii care prezintă mai puține suprapuneri funcționale. Cu toate acestea, chiar dacă BIEN a compilat date voluminoase despre parcele care permit estimări ale abundenței locale, realizarea unor comparații rezonabile între diferite regiuni și forme de creștere este îngreunată de eșantionarea inegală, de diferențele regionale în ceea ce privește diversitatea gamma și de metodele incomensurabile de cuantificare a abundenței.
Planuri comune în cadrul biomurilor
În ciuda lacunelor persistente din datele disponibile, analizele noastre au scos la iveală o relație nedocumentată anterior, comună tuturor biomurilor din emisfera vestică: în orice biom, cele mai răspândite specii tind, de asemenea, să prezinte o distinctivitate funcțională scăzută, în timp ce speciile cele mai distinctive din punct de vedere funcțional sunt aproape invariabil restrânse în distribuția lor (Figura 3). Această relație “ocupație-redundanță” poate sugera că condițiile climatice predominante într-un biom selectează un set de caracteristici comune, speciile mai distincte din punct de vedere funcțional fiind limitate la un subset mai rar de habitate din cadrul biomului. Prevalența generală a speciilor asemănătoare din punct de vedere funcțional în toate biomurile și relația “ocupare-redundanță” sunt ambele în concordanță cu o analiză globală recentă a diversității funcționale la nivel de comunitate care sugerează că filtrarea habitatelor duce la coexistența speciilor asemănătoare din punct de vedere funcțional (Li et al., 2018), precum și cu studiile care arată că redundanțele funcționale cresc stabilitatea comunității (de exemplu, Walker et al., 1999; Pillar et al., 2013). Mai mult, împreună cu gradul ridicat de suprapunere atât taxonomică, cât și funcțională între biomi (figurile 1, 5), faptul că speciile comune și răspândite sunt similare din punct de vedere funcțional întărește ideea că plantele terestre dintr-o gamă largă de condiții de mediu au în comun caracteristici comune aproape de nucleul spectrului de trăsături funcționale (Wright et al., 2004; Díaz et al., 2016). Spre deosebire de Umaña et al. (2017), rezultatele noastre arată că speciile rare și restrânse din punct de vedere geografic pot sau nu să fie distincte din punct de vedere funcțional de speciile mai răspândite și mai comune. Astfel, rămâne deschisă întrebarea dacă speciile mai distincte din punct de vedere funcțional sunt specialiști în anumite medii sau dacă combinațiile lor de trăsături rezultă în compromisuri demografice sau fiziologice care le limitează distribuția geografică.
Comparații între biomi
Compararea noastră a distribuțiilor hipervolumului trăsăturilor între biomi (Figura 2B) este în concordanță cu observația că mediile mai bogate în specii sunt, de asemenea, mai diverse din punct de vedere funcțional (Swenson et al., 2012; Lamanna et al., 2014; Li et al., 2018; Šímová et al., 2018). Rezultatele noastre sunt mai echivoce în ceea ce privește ipoteza conform căreia mediile climatice sezoniere și extreme limitează în mod constant diversitatea funcțională a speciilor (de la Riva et al., 2018). Toate biomurile tropicale prezintă o diversitate funcțională medie mai mare decât toate biomurile temperate, iar biomurile polare prezintă într-adevăr cele mai mici hipervolumuri. Cu toate acestea, în cadrul fiecărui grup, biomii mai secetoși sau mai variabili din punct de vedere sezonier nu afișează întotdeauna hipervolumele mai mici (de exemplu, pădurile uscate, pădurile xerice), așa cum ne-am fi așteptat în urma ipotezei de toleranță a lui Currie et al. (2004), în timp ce condițiile climatice optime susțin mai multe combinații de parametri fiziologici. Doar din aceste rezultate nu putem determina dacă biomurile temperate și polare sunt mai puțin diverse din punct de vedere taxonomic din cauza limitelor spațiului de nișă funcțional sau dacă hipervolumele lor funcționale sunt mici pentru că nu sunt diverse din punct de vedere taxonomic. Abordările de modelare nulă ar putea ajuta, potențial, la disocierea diversității taxonomice și funcționale (Swenson et al., 2012; Lamanna et al., 2014; Šímová et al., 2018), dar o astfel de analiză a depășit sfera de aplicare a acestui studiu. Mai important, rezultatele noastre consolidează importanța înțelegerii modului în care istoria evolutivă și biogeografică modelează diversitatea funcțională a biomurilor (Woodward et al., 2004; Pennington et al., 2006; Donoghue și Edwards, 2014; Moncrieff et al., 2016). Împărtășirea extinsă a speciilor (și a liniilor superioare) între biome în cadrul diferitelor domenii biogeografice, dar, în mare parte, nu și între acestea, ar putea servi atât la omogenizarea diversității funcționale în cadrul domeniilor, cât și la furnizarea de indicii cu privire la trăsăturile caracteristice care sunt selectate pentru sau împotriva lor de diferite medii (Douma et al, 2012; Zanne et al., 2014, 2018).
În ciuda suprapunerii substanțiale a hipervolumului între toate biomurile (Figura suplimentară 2), biomurile tropicale, temperate și reci par să ocupe toate regiuni distincte ale spațiului funcțional (Figurile 5B,C). Principalele trăsături care diferențiază biomurile par a fi trăsături legate de dimensiunea generală a plantelor, inclusiv atât înălțimea la maturitate, cât și masa semințelor (figurile suplimentare 7-10), mai degrabă decât trăsături economice ale frunzelor, așa cum s-a observat în analize mai locale, bazate pe parcele (de exemplu, Douma et al., 2012). Excepție face P din frunze, care a prezentat diferențe substanțiale între biomul tropical și cel temperat/polar (figurile suplimentare 7-10), un model care a fost observat în alte analize bazate atât pe valori specifice fiecărei specii, cât și pe măsurători ale întregului ecosistem (Kerkhoff et al., 2005; Swenson et al., 2012). Deoarece P din frunze a fost trăsătura noastră cea mai puțin eșantionată, diferențele dintre tărâmurile tropicale și cele temperate/polare ar putea fi supuse unor prejudecăți cauzate de procedura de imputare. Cu toate acestea, Leaf P prezintă un semnal filogenetic semnificativ (Kerkhoff et al., 2006), ceea ce sugerează că imputările nu ar trebui să fie părtinitoare (Swenson et al., 2017). Leaf P este, de asemenea, semnificativ mai mare în formele de creștere erbacee (Kerkhoff et al., 2006), astfel încât trecerea latitudinală de la o diversitate predominant lemnoasă la o diversitate erbacee (Figura 4) poate, de asemenea, să influențeze acest model.
Când am limitat analizele hipervolumului doar la speciile cele mai răspândite și cele mai comune din punct de vedere funcțional în fiecare biom, biomurile individuale din cadrul fiecărui tărâm biogeografic s-au suprapus mai puțin în spațiul trăsăturilor funcționale (comparați Figurile 5B,C), ceea ce sugerează că aceste specii pot reflecta fenotipuri mai bine adaptate la anumite medii. În acest context, biomul pădurilor xerice este deosebit de interesant. În parte datorită includerii ecoregiunii Chaco, pădurile xerice se grupează cu biomurile tropicale din punct de vedere taxonomic (Figura 5A). Însă, atunci când sunt luate în considerare hipervolumele funcționale ale tuturor speciilor, acestea prezintă o asemănare mult mai puternică cu biomurile temperate (Figura 5B). În cele din urmă, atunci când sunt analizate doar speciile cele mai răspândite și cele mai comune din punct de vedere funcțional, pădurile xerice prezintă din nou o similitudine sporită cu biomii tropicali, dar, de asemenea, mențin o similitudine ridicată cu biomii temperați de pășuni și mediteraneeni (Figura 5C). Tranzițiile de la mediile calde, mesice la mediile mai reci, mai uscate și mai sezoniere sunt facilitate de trăsături similare, de exemplu, obiceiul erbaceu (Douma et al., 2012; Zanne et al., 2014, 2018) și, la fel ca mediile mai reci, comunitățile din mediile uscate tind, de asemenea, să fie mai grupate filogenetic (Qian și Sandel, 2017). În plus, poziția zonelor xerice la limita zonei de convergență intertropicale face ca acestea să reprezinte o tranziție geografică între tărâmul tropical și cel temperat. Faptul că pădurile xerice sunt intermediare între tărâmul tropical și cel temperat, atât din punct de vedere funcțional, cât și biogeografic, întărește și mai mult ideea că, pentru a înțelege mai bine diversitatea funcțională a biomurilor, trebuie să luăm în considerare istoricul biogeografic și filogenetic al acestora (Pennington et al., 2006; Donoghue și Edwards, 2014; Moncrieff et al, 2016).
Concluzii
Orice clasificare a biomurilor terestre impune un număr mic de categorii discrete pe gradiente continue în ceea ce privește clima și distribuția speciilor și, prin urmare, reprezintă o simplificare grosolană a peisajului ecologic complex (Moncrieff et al., 2016, dar a se vedea Silva de Miranda et al., 2018). Cu toate acestea, în ciuda potențialului lor de simplificare excesivă, biomii sunt construcții utile pentru organizarea și înțelegerea biodiversității și funcționării principalelor ecosisteme terestre ale Pământului, iar abordările bazate pe trăsături au un potențial ridicat de a ajuta la modelarea dinamică a distribuției globale a vegetației.
În acest studiu am arătat că o mare parte din diversitatea taxonomică a tuturor biomilor reprezintă specii care sunt atât distribuite în mod îngust, cât și similare din punct de vedere funcțional. Mai mult, în cadrul biomurilor, speciile cele mai distinctive din punct de vedere funcțional din fiecare biom tind, de asemenea, să fie rare din punct de vedere geografic, în timp ce speciile răspândite prezintă în mod uniform o distinctivitate funcțională scăzută. În ciuda suprapunerii taxonomice și funcționale extinse între biomi, aceștia se grupează în unități distincte, distincte din punct de vedere biogeografic și climatic, în special atunci când gruparea funcțională se bazează pe speciile cele mai răspândite și comune din punct de vedere funcțional din fiecare biom. Cu toate acestea, avansarea unei înțelegeri funcționale a biomurilor va necesita nu doar o mai bună caracterizare a variației trăsăturilor în cadrul și între tipurile de vegetație, ci și informații privind istoria biogeografică și filogenetică a ansamblurilor de specii și abundența relativă a speciilor în cadrul biomurilor.
Date disponibile
Seturile de date privind trăsăturile și hărțile de distribuție a speciilor analizate pentru acest studiu pot fi descărcate prin intermediul pachetului R BIEN (Maitner et al., 2018). A se vedea http://bien.nceas.ucsb.edu/bien/ pentru mai multe detalii despre baza de date BIEN.
Contribuții ale autorilor
SE-L, AK, BJE, DMN și CV au conceput studiul; SE-L și AK au analizat datele; SE-L și AK au scris prima versiune a manuscrisului; CV, BB, NK, BSM, BM, RKP, BS, BS, SS, J-CS și SKW au contribuit la colaționarea și crearea bazei de date cu trăsături și hărți de distribuție. Toți autorii au contribuit la elaborarea și redactarea manuscrisului.
Finanțare
SE-L, DMN și AK au fost susținuți de un grant de cercetare în colaborare de la US National Science Foundation (DEB-1556651). BJE a fost sprijinit de grantul Fundației Naționale pentru Știință DEB-1457812 și Macrosystems-1065861. CV a fost sprijinit de Consiliul European de Cercetare (grantul ERC-StG-2014-639706-CONSTRAINTS) și de Fundația Franceză pentru Cercetarea Biodiversității (FRB; www.fondationbiodiversite.fr) în contextul proiectului CESAB Cauze și consecințe ale rarității funcționale de la scară locală la scară globală. J-CS consideră că această lucrare este o contribuție la proiectul său VILLUM Investigator Biodiversity Dynamics in a Changing World, finanțat de VILLUM FONDEN (grant 16549). SKW a fost sprijinit de Strategic Science Investment Fund al New Zealand Ministry of Business, Innovation and Employment’s Science and Innovation Group din cadrul New Zealand Ministry of Business, Innovation and Employment.
Declarație privind conflictul de interese
Autorii declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
Recunoștințe
Îi mulțumim lui Dan Griffith și celor doi recenzenți pentru critica lor constructivă care a îmbunătățit foarte mult acest manuscris. Mulțumim, de asemenea, tuturor contribuitorilor de date BIEN (a se vedea http://bien.nceas.ucsb.edu/bien/data-contributors/ pentru o listă completă).
Supplementary Material
Materialul suplimentar pentru acest articol poate fi găsit online la adresa: https://www.frontiersin.org/articles/10.3389/fevo.2018.00219/full#supplementary-material
Banda, R.K., Delgado-Salinas, A., Dexter, K. G., Linares-Palomino, R., Oliveira-Filho, A., Prado, D., et al. (2016). Modele de diversitate a plantelor în pădurile uscate neotropicale și implicațiile lor pentru conservare. Science 353, 1383-1387. doi: 10.1126/science.aaf5080
PubMed Abstract | Full CrossRef Text | Google Scholar
Blonder, B., Lamanna, C., Violle, C., și Enquist, B. J. (2014). Hipervolumul n-dimensional. Glob. Ecol. Biogeogr. 23, 595-609. doi: 10.1111/geb.12146
CrossRef Full Text | Google Scholar
Blonder, B., Morrow, C. B., Maitner, B., Harris, D. J., Lamanna, C., Violle, C., et al. (2018). Noi abordări pentru delimitarea hipervolumelor n-dimensionale. Methods Ecol. Evol. 9, 305-319. doi: 10.1111/2041-210X.12865
CrossRef Full Text | Google Scholar
Bonan, G. B., Oleson, K. W., Fisher, R. A., Lasslop, G., și Reichstein, M. (2012). Reconcilierea trăsăturilor fiziologice ale frunzelor și a datelor de flux din coronament: utilizarea bazelor de date TRY și FLUXNET în Community Land Model versiunea 4. J. Geophys. Res. Biogeosci. 117:G02026. doi: 10.1029/2011JG001913
CrossRef Full Text | Google Scholar
Borregaard, M. K., și Rahbek, C. (2010). Cauzalitatea relației dintre distribuția geografică și abundența speciilor. Q. Rev. Biol. 85, 3-25. doi: 10.1086/650265
PubMed Abstract | Full CrossRef Text | Google Scholar
Bruggeman, J., Heringa, J., și Brandt, B. W. (2009). PhyloPars: estimarea valorilor parametrilor lipsă folosind filogenia. Nucleic Acids Res. 37, W179-W184. doi: 10.1093/nar/gkp370
PubMed Abstract | CrossRef Full Text | Google Scholar
Cadotte, M. W., Carscadden, K., și Mirotchnick, N. (2011). Dincolo de specii: diversitatea funcțională și menținerea proceselor și serviciilor ecologice. J. Appl. Ecol. 48, 1079-1087. doi: 10.1111/j.1365-2664.2011.02048.x
CrossRef Full Text | Google Scholar
Cornwell, W. K., Cornelissen, J. H., Amatangelo, K., Dorrepaal, E., Eviner, V. T., Godoy, O., et al. (2008). Caracteristicile speciilor de plante reprezintă controlul predominant asupra ratelor de descompunere a litierei în cadrul biomurilor din întreaga lume. Ecol. Lett. 11, 1065-1071. doi: 10.1111/j.1461-0248.2008.01219.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Craine, J. M. (2009). Strategii de resurse ale plantelor sălbatice. New Jersey, NJ: Princeton University Press. doi: 10.1515/9781400830640
CrossRef Full Text | Google Scholar
Currie, D. J., Mittelbach, G. G., Cornell, H. V., Field, R., Guégan, J. F., Hawkins, B. A., et al. (2004). Predicții și teste ale ipotezelor bazate pe climă privind variația la scară largă a bogăției taxonomice. Ecol. Lett. 12, 1121-1134. doi: 10.1111/j.1461-0248.2004.00671.x
CrossRef Full Text | Google Scholar
de la Riva, E. G., Violle, C., Pérez-Ramos, I. M., Marañón, T., Navarro-Fernández, C. M., Olmo, M., et al. (2018). O abordare multidimensională a trăsăturilor funcționale dezvăluie amprenta stresului de mediu în comunitățile lemnoase mediteraneene. Ecosystems 21, 248-262. doi: 10.1007/s10021-017-0147-7
CrossRef Full Text | Google Scholar
Díaz, S., Kattge, J., Cornelissen, J. H., Wright, I. J., Lavorel, S., Dray, S., et al. (2016). The global spectrum of plant form and function (Spectrul global al formei și funcției plantelor). Nature 529, 167-171. doi: 10.1038/nature16489
PubMed Abstract | Text integral | Google Scholar
Díaz, S., Lavorel, S., de Bello, F., Quétier, F., Grigulis, K., și Robson, T. M. (2007). Încorporarea efectelor diversității funcționale a plantelor în evaluările serviciilor ecosistemice. Proc. Natl. acad. Sci. U.S.A 104, 20684-20689. doi: 10.1073/pnas.0704716104
PubMed Abstract | CrossRef Full Text | Google Scholar
Donoghue, M. J., și Edwards, E. J. (2014). Schimbările biomei și evoluția nișei la plante. Annu. Rev. Ecol. Evol. sist. sist. 45, 547-572. doi: 10.1146/annurev-ecolsys-120213-091905
CrossRef Full Text | Google Scholar
Douma, J. C., Aerts, R., Witte, J. P. M. M., Bekker, R. M., Kunzmann, D., Metselaar, K., et al. (2012). O combinație de trăsături vegetale diferite din punct de vedere funcțional oferă un mijloc de a prezice cantitativ o gamă largă de ansambluri de specii în nord-vestul Europei. Ecography 35, 364-373. doi: 10.1111/j.1600-0587.2011.07068.x
CrossRef Full Text | Google Scholar
Engemann, K., Sandel, B., Enquist, B. J., Jørgensen, P. M., Kraft, N., Marcuse-Kubitza, A., et al. (2016). Patterns and drivers of plant functional group dominance across the Western Hemisphere: a macroecological re-assessment based on a massive botanical dataset. Bot. J. Linn. Soc. 180, 141-160. doi: 10.1111/boj.12362
CrossRef Full Text | Google Scholar
Enquist, B. J., Condit, R., Peet, R., Thiers, B., și Schildhauer, M. (2016). Rețeaua de informații botanice și ecologie (BIEN): Infrastructură cibernetică pentru o rețea integrată de informații botanice pentru a investiga impactul ecologic al schimbărilor climatice globale asupra biodiversității plantelor. PeerJ Prepr. 4:e2615v2. doi: 10.7287/peerj.preprints.2615v2
CrossRef Full Text | Google Scholar
Garnier, E., Cortez, J., Billès, G., Navas, M. L., Roumet, C., Debussche, M., et al. (2004). Markerii funcționali ai plantelor captează proprietățile ecosistemului în timpul succesiunii secundare. Ecology 85, 2630-2637. doi: 10.1890/03-0799
CrossRef Full Text | Google Scholar
Goldsmith, G. R., Morueta-Holme, N., Sandel, B., Fitz, E. D., Fitz, S. D., Boyle, B., et al. (2016). Plant-O-Matic: un ghid dinamic și mobil pentru toate plantele din cele două Americi. Methods Ecol. Evol. 7, 960-965. doi: 10.1111/2041-210X.12548
CrossRef Full Text | Google Scholar
Goolsby, E. W., Bruggeman, J., și Ané, C. (2017). Rphylopars: metode comparative filogenetice multivariate rapide pentru date lipsă și variație în cadrul speciilor. Methods Ecol. Evol. 8, 22-27. doi: 10.1111/2041-210X.12612
CrossRef Full Text | Google Scholar
Grenié, M., Denelle, P., Tucker, C. M., Munoz, F., și Violle, C. (2017). funrar: un pachet R pentru caracterizarea rarității funcționale. Divers. Distrib. 23, 1365-1371. doi: 10.1111/ddi.12629
CrossRef Full Text | Google Scholar
Hortal, J., de Bello, F., Diniz-Filho, J. A. F., J. A. F., Lewinsohn, T. M. M., Lobo, J. M., și Ladle, R. J. (2015). Șapte deficiențe care afectează cunoașterea la scară largă a biodiversității. Annu. Rev. Ecol. Evol. Syst. 46, 523-549. doi: 10.1146/annurev-ecolsys-112414-054400
CrossRef Full Text | Google Scholar
Kerkhoff, A. J., Enquist, B. J., Elser, J. J., și Fagan, W. F. (2005). Plant allometry, stoichiometry and the temperature-dependence of primary productivity. Glob. Ecol. Biogeogr. 14, 585-598. doi: 10.1111/j.1466-822X.2005.00187.x
CrossRef Full Text | Google Scholar
Kerkhoff, A. J., Fagan, W. F. F., Elser, J. J., și Enquist, B. J. (2006). Variația filogenetică și a formei de creștere în scalarea azotului și a fosforului în plantele de semințe. Am. Nat. 168, E103-E122. doi: 10.1086/507879
PubMed Abstract |Ref Full Text | Google Scholar
Kraft, N. J., Valencia, R. și Ackerly, D. D. (2008). Trăsături funcționale și asamblarea comunității de arbori bazată pe nișă într-o pădure amazoniană. Science 322, 580-582. doi: 10.1126/science.1160662
PubMed Abstract | Text integral | Google Scholar
Lamanna, C., Blonder, B., Violle, C., Kraft, N. J., Sandel, B., et al. (2014). Functional trait space and the latitudinal diversity gradient. Proc. Natl. acad. Sci. U.S.A. 111, 13745-13750. doi: 10.1073/pnas.1317722111
PubMed Abstract | CrossRef Full Text | Google Scholar
Lavorel, S., și Garnier, E. (2002). Predicția schimbărilor în compoziția comunității și funcționarea ecosistemului din trăsăturile plantelor: revizitarea Sfântului Graal. Funct. Ecol. 16, 545-556. doi: 10.1046/j.1365-2435.2002.00664.x
CrossRef Full Text | Google Scholar
Lavorel, S., Grigulis, K., Lamarque, P., Colace, M.-P., Garden, D., Girel, J., et al. (2011). Utilizarea trăsăturilor funcționale ale plantelor pentru a înțelege distribuția peisagistică a mai multor servicii ecosistemice. J. Ecol. 99, 135-147. doi: 10.1111/j.1365-2745.2010.01753.x
CrossRef Full Text | Google Scholar
Li, Y., Shipley, B., Price, J. N., Dantas, V. de L., Tamme, R., Westoby, M., et al. (2018). Filtrarea habitatelor determină ocuparea nișei funcționale a comunităților de plante din întreaga lume. J. Ecol. 106, 1001-1009. doi: 10.1111/1365-2745.12802
CrossRef Full Text | Google Scholar
Magallón, S., Gómez-Acevedo, S., Sánchez-Reyes, L. L., și Hernández-Hernández, T. (2015). Un arbore temporal metacalibrat documentează creșterea timpurie a diversității filogenetice a plantelor cu flori. New Phytol. 207, 437-453. doi: 10.1111/nph.13264
PubMed Abstract | Textul integral | Google Scholar
Maitner, B. S., Boyle, B., Casler, N., Condit, R., Donoghue, J., Durán, S. M., et al. (2018). Pachetul BIEN R: un instrument de accesare a bazei de date a rețelei de informații botanice și ecologie (BIEN). Methods Ecol. Evol. 9, 373-379. doi: 10.1111/2041-210X.12861
CrossRef Full Text | Google Scholar
Moncrieff, G. R., Bond, W. J., și Higgins, S. I. (2016). Revizuirea conceptului de biom pentru înțelegerea și predicția impactului schimbărilor globale. J. Biogeogr. 43, 863-873. doi: 10.1111/jbi.12701
CrossRef Full Text | Google Scholar
Novosolov, M., Rodda, G. H., North, A. C., Butchart, S. H. M., Tallowin, O. J. S. S., Gainsbury, A. M., et al. (2017). Relația dintre densitatea populației și mărimea razei de acțiune a populației revizuită. Glob. Ecol. Biogeogr. 26, 1088-1097. doi: 10.1111/geb.12617
CrossRef Full Text | Google Scholar
Olson, D. M., Dinerstein, E., Wikramanayake, E. D., Burgess, N. D., Powell, G. V. N., Underwood, E. C., et al. (2001). Terrestrial Ecoregions of the world: a new map of life on earth: a new global map of terrestrial ecoregions provides an innovative tool for conserving biodiversity (Ecoregiunile terestre ale lumii: o nouă hartă a vieții pe pământ: o nouă hartă globală a ecoregiunilor terestre oferă un instrument inovator pentru conservarea biodiversității). Bioscience 51, 933-938. doi: 10.1641/0006-3568(2001)0512.0.CO;2
CrossRef Full Text | Google Scholar
Pennington, R. T., Prado, D. E., și Pendry, C. A. (2000). Neotropical seasonally dry forests and quaternary vegetation changes. J. Biogeogr. 27, 261-273. doi: 10.1046/j.1365-2699.2000.00397.x
CrossRef Full Text | Google Scholar
Pennington, R. T., Richardson, J. E., și Lavin, M. (2006). Insights into the historical construction of species-rich biomes from dated plant phylogenies, neutral ecological theory and phylogenetic community structure. New Phytol. 172, 605-616. doi: 10.1111/j.1469-8137.2006.01902.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Penone, C., Davidson, A. D., Shoemaker, K. T., Di Marco, M., Rondinini, C., Brooks, T. M., et al. (2014). Imputarea datelor lipsă în seturile de date privind trăsăturile istoriei vieții: ce abordare are cea mai bună performanță? Methods Ecol. Evol. 5, 961-970. doi: 10.1111/2041-210X.12232
CrossRef Full Text | Google Scholar
Pillar, V. D., Blanco, C. C., Müller, S. C., Sosinski, E. E., Joner, F., și Duarte, L. D. S. (2013). Redundanța funcțională și stabilitatea în comunitățile de plante. J. Veg. Sci. 24, 963-974. doi: 10.1111/jvs.12047
CrossRef Full Text | Google Scholar
Prado, D. E., și Gibbs, P. E. (1993). Patterns of species distributions in the dry seasonal forests of South America. Ann. Mo. Bot. Gard. 80, 902-927. doi: 10.2307/2399937
CrossRef Full Text | Google Scholar
Qian, H., și Sandel, B. (2017). Structura filogenetică a ansamblurilor regionale de angiosperme de-a lungul gradienților latitudinali și climatici din America de Nord. Glob. Ecol. Biogeogr. 26, 1258-1269. doi: 10.1111/geb.12634
CrossRef Full Text | Google Scholar
Reich, P. B., Wright, I. J., Cavender-Bares, J., Craine, J. M., Oleksyn, J., Westoby, M., et al. (2003). Evoluția variației funcționale a plantelor: trăsături, spectre și strategii. Int. J. Plant Sci. 164, S143-S164. doi: 10.1086/374368
CrossRef Full Text | Google Scholar
Silva de Miranda, P. L., Oliveira-Filho, A., Pennington, R. T., Neves, D. M., Baker, T. R., și Dexter, K. G. (2018). Utilizarea inventarelor de specii de arbori pentru a cartografia biomurile și a evalua suprapunerile climatice ale acestora în America de Sud tropicală de câmpie. Glob. Ecol. Biogeogr. 27, 899-912. doi: 10.1111/geb.12749
CrossRef Full Text | Google Scholar
Šímová, I., Violle, C., Kraft, N. J. B., Storch, D., Svenning, J.-C., Boyle, B., et al. (2015). Modificări ale mediilor și varianțelor trăsăturilor în ansamblurile de arbori din America de Nord: modelele de bogăție a speciilor sunt slab legate de spațiul funcțional. Ecography 38, 649-658. doi: 10.1111/ecog.00867
CrossRef Full Text | Google Scholar
Šímová, I., Violle, C., Svenning, J.-C., Kattge, J., Engemann, K., Sandel, B., et al. (2018). Modelele spațiale și relațiile climatice ale principalelor trăsături ale plantelor din Lumea Nouă diferă între speciile lemnoase și erbacee. J. Biogeogr. 45, 895-916. doi: 10.1111/jbi.13171
CrossRef Full Text | Google Scholar
Smith, S. A., și Beaulieu, J. M. (2009). Istoria vieții influențează ratele de evoluție a nișei climatice la plantele cu flori. Proc. Biol. Sci. 276, 4345-4352. doi: 10.1098/rspbb.2009.1176
PubMed Abstract | Textul integral | Google Scholar
Smith, S. A., și Brown, J. W. (2018). Construirea unei filogenii de plante de semințe larg incluzive. Am. J. Bot. 105, 302-314. doi: 10.1002/ajb2.1019
PubMed Abstract | Full Text | Google Scholar
Smith, S. A., și Donoghue, M. J. (2008). Ratele de evoluție moleculară sunt legate de istoria vieții la plantele cu flori. Science 322, 86-89. doi: 10.1126/science.1163197
PubMed Abstract | Textul integral | Google Scholar
Swenson, N. G. (2014). Imputarea filogenetică a bazelor de date de trăsături funcționale ale plantelor. ecography 37, 105-110. doi: 10.1111/j.1600-0587.2013.00528.x
CrossRef Full Text
Swenson, N. G., Enquist, B. J., Pither, J., Kerkhoff, A. J., Boyle, B., Weiser, M. D., et al. (2012). Biogeografia și filtrarea diversității funcționale a plantelor lemnoase din America de Nord și de Sud. Glob. Ecol. Biogeogr. 21, 798-808. doi: 10.1111/j.1466-8238.2011.00727.x
CrossRef Full Text | Google Scholar
Swenson, N. G., Weiser, M. D., Mao, L., Araújo, M. B., Diniz-Filho, J. A. F., Kollmann, J., et al. (2017). Filogenia și predicția diversității funcționale a arborilor în medii continentale noi. Glob. Ecol. Biogeogr. 26, 553-562. doi: 10.1111/geb.12559
CrossRef Full Text | Google Scholar
Umaña, M. N., Mi, X., Cao, M., Enquist, B. J., Hao, Z., Howe, R., et al. (2017). Rolul unicității funcționale și al agregării spațiale în explicarea rarității în arbori. Glob. Ecol. Biogeogr. 26, 777-786. doi: 10.1111/geb.12583
CrossRef Full Text | Google Scholar
van Bodegom, P. M., Douma, J. C., și Verheijen, L. M. (2014). O abordare complet bazată pe trăsături pentru modelarea distribuției globale a vegetației. Proc. Natl. acad. Sci. U.S.A. 111, 13733-13738. doi: 10.1073/pnas.1304551110
PubMed Abstract | CrossRef Full Text | Google Scholar
Violle, C., Reich, P. B., Pacala, S. W., Enquist, B. J., și Kattge, J. (2014). The emergence and promise of functional biogeography (Apariția și promisiunea biogeografiei funcționale). Proc. Natl. acad. Sci. U.S.A. 111, 13690-13696. doi: 10.1073/pnas.1415442111
PubMed Abstract | CrossRef Full Text | Google Scholar
Violle, C., Thuiller, W., Mouquet, N., Munoz, F., Kraft, N. J. B., Cadotte, M. W., et al. (2017). Raritatea funcțională: ecologia valorilor aberante. Trends Ecol. Evol. 32, 356-367. doi: 10.1016/j.tree.2017.02.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Walker, B., Kinzig, A., și Langridge, J. (1999). Diversitatea atributelor plantelor, reziliența și funcția ecosistemului: natura și semnificația speciilor dominante și minore. Ecosystems 2, 95-113. doi: 10.1007/s100219900062
CrossRef Full Text | Google Scholar
Westoby, M. (1998). O schemă de strategie de ecologie vegetală a înălțimii frunzelor-semințe (LHS). Plant Soil 199, 213-227. doi: 10.1023/A:1004327224729
CrossRef Full Text | Google Scholar
Woodward, F. I., Lomas, M. R., and Kelly, C. K. (2004). Clima globală și distribuția biomurilor vegetale. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 359, 1465-1476. doi: 10.1098/rstb.2004.1525
PubMed Abstract | CrossRef Full Text | Google Scholar
Wright, I. J., Reich, P. B. B., Westoby, M., Ackerly, D. D., Baruch, Z., Bongers, F., et al. (2004). The worldwide leaf economics spectrum. Nature 428, 821-827. doi: 10.1038/nature02403
PubMed Abstract | Textul integral | Google Scholar
Xia, J., Niu, S., Ciais, P., Janssens, I. A., Chen, J., Ammann, C., et al. (2015). Controlul comun al productivității primare brute terestre brute de către fenologia și fiziologia plantelor. Proc. Natl. acad. Sci. U.S.A. 112, 2788-2793. doi: 10.1073/pnas.1413090112
PubMed Abstract | CrossRef Full Text | Google Scholar
Zanne, A. E., Pearse, W. D., Cornwell, W. K., McGlinn, D. J., Wright, I. J., și Uyeda, J. C. (2018). Biogeografia funcțională a angiospermelor: viața la extreme. New Phytol. 218, 1697-1709. doi: 10.1111/nph.15114
PubMed Abstract | CrossRef Full Text | Google Scholar
Zanne, A. E., Tank, D. C., Cornwell, W. K., Eastman, J. M., Smith, S. A., FitzJohn, R. G., et al. (2014). Trei chei pentru radiația angiospermelor în medii înghețate. Nature 506, 89-92. doi: 10.1038/nature12872
PubMed Abstract | Refef Full Text | Google Scholar
.