Evaluare | Biopsihologie | Comparată |Cognitivă | Dezvoltare | Limbaj | Diferențe individuale |Personalitate | Filosofie | Socială |
Metode | Statistică |Clinică | Educațională | Industrială | Articole profesionale | Psihologie mondială |
Biologică:Genetică comportamentală – Psihologie evolutivă – Neuroanatomie – Neurochimie – Neuroendocrinologie – Neuroștiințe – Psihoneuroimunologie – Psihologie fiziologică – Psihofarmacologie(Index, Schiță)
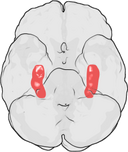
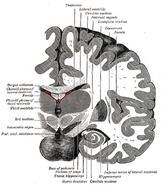
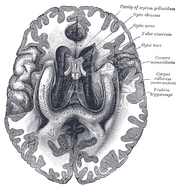
Localizarea hipocampusului în creierul uman.
Pentru revista cu același nume, vezi Hipocampus (revistă)
Hipocampul este o parte a creierului anterior, situat în lobul temporal medial. Acesta aparține sistemului limbic și joacă roluri majore în memoria pe termen lung și în navigația spațială. Oamenii și alte mamifere au două hipocampi, câte unul în fiecare parte a creierului. La rozătoare, unde a fost studiat cel mai mult, hipocampul are o formă asemănătoare unei banane. La oameni, acesta are o formă curbată și întortocheată care le-a amintit primilor anatomiști de un căluț de mare. Numele, de fapt, derivă din cuvântul grecesc pentru căluț de mare (greacă: hippos = cal, campos = mare).
În boala Alzheimer, hipocampul este una dintre primele regiuni ale creierului care suferă leziuni; problemele de memorie și dezorientarea apar printre primele simptome. Afectarea hipocampusului poate rezulta și în urma lipsei de oxigen (anoxie), a encefalitei sau a epilepsiei lobului temporal mezial. Persoanele cu leziuni extinse ale hipocampului pot suferi amnezie, adică incapacitatea de a forma sau de a reține noi amintiri.
Funcțiile hipocampusului
Poate că cea mai veche idee a fost aceea că hipocampul este implicat în olfacție: acest lucru pare să fi fost sugerat în principal de localizarea sa în creier, lângă cortexul olfactiv. Continuă să existe un oarecare interes pentru răspunsurile olfactive ale hipocampului, dar aproape nimeni nu mai crede acum că funcția primară a hipocampului este olfactivă.
De-a lungul anilor, trei idei principale despre funcția hipocampului au dominat literatura de specialitate: inhibiția, memoria și spațiul. Teoria inhibiției comportamentale (caricaturizată de O’Keefe și Nadel ca “apasă pe frâne!”) a fost foarte populară până în anii 1960. Ea și-a derivat o mare parte din forța sa din două observații: în primul rând, animalele cu leziuni ale hipocampului tind să fie hiperactive; în al doilea rând, animalele cu leziuni ale hipocampului au adesea dificultăți în a învăța să inhibe răspunsurile pe care le-au învățat anterior. Jeffrey Gray a dezvoltat această linie de gândire într-o teorie cu drepturi depline a rolului hipocampusului în anxietate. Cu toate acestea, teoria inhibiției nu este foarte populară în prezent.
A doua linie de gândire importantă leagă hipocampul de memorie. Deși a avut precursori, această idee și-a derivat forța principală dintr-un raport foarte cunoscut al lui Scoville și Milner despre rezultatele distrugerii chirurgicale a hipocampusului (în încercarea de a ameliora crizele epileptice), la un pacient cunoscut sub numele de H.M. Rezultatul neașteptat a fost o amnezie severă: H.M. a fost incapabil să-și amintească în mod conștient evenimentele care au avut loc după operație sau timp de câțiva ani înainte de aceasta. Acest caz a suscitat un interes atât de mare încât se spune acum că H.M. este cel mai intens studiat caz medical din istorie. În anii care au urmat, au fost studiați și alți pacienți cu niveluri similare de afectare a hipocampului și amnezie (cauzată de accidente sau boli) și literalmente mii de experimente au studiat fiziologia plasticității neuronale în hipocampus. În prezent, există un acord aproape universal asupra faptului că hipocampul joacă un fel de rol important în memorie; cu toate acestea, natura exactă a acestui rol rămâne larg dezbătută.
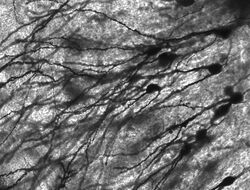
Neuroni colorați de Golgi în hipocampul rozătoarelor.
Rolul în memorie
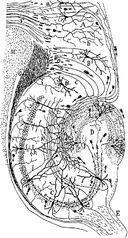
Desenare a circuitului neuronal al hipocampusului rozătoarelor. S. Ramón y Cajal, 1911.
Psihologii și neuroștiințele contestă rolul precis al hipocampusului, dar în general sunt de acord că acesta are un rol esențial în formarea de noi amintiri despre evenimente trăite personal (memorie episodică sau autobiografică). Unii cercetători preferă să considere hipocampul ca făcând parte dintr-un sistem de memorie mai larg, al lobului temporal medial, responsabil pentru memoria declarativă generală (amintiri care pot fi verbalizate în mod explicit – acestea ar include, de exemplu, memoria pentru fapte, pe lângă memoria episodică).
Articol principal: Rolul hipocampusului în memorie
Câteva dovezi implică hipocampul în stocarea și procesarea informațiilor spațiale. Studiile la șobolani au arătat că neuronii din hipocampus au câmpuri de tragere spațiale. Aceste celule se numesc celule de loc. Unele celule se declanșează atunci când animalul se află într-o anumită locație, indiferent de direcția de deplasare, în timp ce majoritatea sunt cel puțin parțial sensibile la direcția capului și la direcția de deplasare.
Articol principal: Rolul hipocampusului în memoria spațială și navigație
Hipocampul și depresia
- Pierderea volumului hipocampal, reflectând poate efectele glucocorticoizilor excesivi asupra
neurogenezei, a fost identificată la grupuri de persoane depresive în comparație cu persoanele de control.
Articol principal: Hipocampul și depresia: Depresia și hipocampul
Istoric
Anatomistul Giulio Cesare Aranzi (circa 1564) a folosit pentru prima dată termenul de hipocampus pentru a descrie organul cerebral datorită asemănării sale vizuale cu un căluț de mare. Acest organ a fost inițial legat de simțul mirosului, mai degrabă decât de funcția sa cunoscută în dobândirea memoriei. Rusul Vladimir Bekhterev a remarcat rolul hipocampusului în memorie în jurul anului 1900, pe baza observațiilor unui pacient cu tulburări profunde de memorie. Cu toate acestea, timp de mulți ani, viziunea convențională asupra hipocampusului a fost că, la fel ca restul sistemului limbic, acesta era responsabil pentru emoții.
Importanța hipocampusului în memorie a fost adusă în atenția cercetătorilor de către pacientul HM. HM suferea de o serie de deficiențe de memorie anterogradă și retrogradă gradată temporal (astfel de deficiențe sunt subiectul filmului Memento) în urma îndepărtării bilaterale a diferitelor structuri ale lobului medio-temporal (inclusiv ablația bilaterală a hipocampusului său) pentru a ameliora crizele epileptice frecvente. De o importanță deosebită este faptul că HM era încă capabil să învețe sarcini procedurale (care sunt asociate cu striatum) și avea un IQ peste medie. HM a demonstrat o disociere unică izbitoare între inteligență și memoria declarativă. Dimensiunea relativă a formațiunii hipocampale în raport cu volumul total al creierului este adesea conservată la majoritatea speciilor de mamifere. Cu toate acestea, s-a constatat că aceste zone sunt relativ hipotrofe la cetacee.
Anatomie
Articol principal: Anatomia hipocampusului
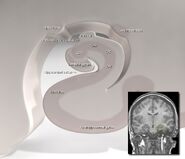
Human hippocampus.
Secțiune coronală colorată cu Nissl a creierului unei maimuțe macaque, arătând hipocampul (încercuit). Sursa: brainmaps.org
Anatomic, hipocampul este o elaborare a marginii cortexului. El poate fi distins ca o zonă în care cortexul se îngustează într-un singur strat de neuroni foartedens, care se încolăcește în formă de S strâns. Structurile care mărginesc marginea cortexului alcătuiesc așa-numitul sistem limbic (din latină limbus = frontieră): acestea includ hipocampul, cortexul cingular, cortexul olfactiv și amigdala. Paul MacLean a sugerat cândva, ca parte a teoriei sale despre creierul trinitar, că structurile limbice cuprind baza neuronală a emoțiilor. Cei mai mulți neuroștiințifici nu mai cred însă că este valabil conceptul de “sistem limbic” unificat.
Hipocampul, în ansamblu, sfârșește prin a arăta ca un fel de tub curbat, care a fostanalogizat în diverse moduri cu un căluț de mare, sau cu un corn de berbec (Cornu Ammonis), sau cu o banană.Această configurație generală este valabilă pentru întreaga gamă de specii de mamifere, de la arici la om, deși detaliile variază. La șobolan, cele două hipocampi seamănă uimitor de mult cu o pereche de banane, unite la tulpină. În creierele umane sau de maimuță, porțiunea de hipocampus din partea de jos, aproape de baza lobului temporal, este mult mai largă decât partea de sus. Una dintre consecințele acestei geometrii complexe este că secțiunile transversale prin hipocampus pot prezenta o varietate deconcertantă de forme, în funcție de unghiul și locația tăieturii.
Cele mai puternice conexiuni ale hipocampusului sunt cu cortexul entorinal (EC), care se află lângă el în lobul temporal. Straturile superficiale ale CE furnizează cele mai numeroase intrări către hipocampus, iar straturile profunde ale CE primesc cele mai numeroase ieșiri. CE, la rândul său, este puternic și reciproc conectat cu multe alte părți ale cortexului. Hipocampul primește, de asemenea, o proiecție foarte importantă din partea zonei septale mediale. Distrugerea zonei septale abolește ritmul theta al hipocampului și afectează grav anumite tipuri de memorie. (Se crede că așa-numitele droguri “date rape” își exercită efectele amnezice cel puțin parțial prin antagonizarea proiecției colinergice de la septul medial la hipocampus.)
Fiziologie
Exemple de EEG al hipocampului de șobolan și de activitate neuronală CA1 în modurile beta (treaz/conduită) și LIA (somn cu unde lente). Fiecare grafic prezintă 20 de secunde de date, cu o urmă EEG hipocampală în partea de sus, rastere de vârfuri de la 40 de celule piramidale CA1 înregistrate simultan în mijloc (fiecare linie de raster reprezintă o celulă diferită) și un grafic al vitezei de alergare în partea de jos. Graficul de sus reprezintă o perioadă de timp în care șobolanul a căutat în mod activ pelete de hrană împrăștiate. Pentru graficul de jos, șobolanul dormea.
Hipocampul prezintă două “moduri” majore de activitate, fiecare asociat cu un model distinct de unde EEG și de activitate a populației neuronale.Aceste moduri sunt denumite după modelele EEG asociate cu ele:theta și activitatea neregulată mare (LIA). Iată câteva dintre principalele lor caracteristici la șobolan, animalul care a fost cel mai mult studiat:
Modul theta apare în timpul stărilor de comportament activ, alert (în special locomoție) și, de asemenea, în timpul somnului REM (visare). În modul theta, EEG-ul este dominat de valuri mari și regulate cu o frecvență de 6-9 Hz, iar principalele grupuri de neuroni hipocampali (celule piramidale și celule granulare) prezintă o activitate de populație rară, ceea ce înseamnă că, în orice interval scurt de timp, marea majoritate a celulelor sunt tăcute, în timp ce mica fracțiune rămasă se declanșează la viteze relativ mari, de până la 50 de vârfuri într-o secundă pentru cele mai active dintre ele. O celulă activă rămâne de obicei activă timp de la o jumătate de secundă la câteva secunde. Pe măsură ce șobolanul se comportă, celulele active devin silențioase și noi celule devin active, dar procentul total de celule active rămâne mai mult sau mai puțin constant. În multe situații, celactivitatea celulară este determinată în mare măsură de localizarea spațială a animalului,dar și alte variabile comportamentale o influențează în mod clar.
Modul LIA apare în timpul somnului cu unde lente (care nu visează), dar șiîn timpul stărilor de imobilitate în stare de veghe, cum ar fi odihna sau mâncatul. În modulLIA, EEG-ul este dominat de unde ascuțite, care sunt deviații mari ale semnalului EEG, sincronizate aleatoriu, cu o durată de 200-300msec. Aceste unde ascuțite determină, de asemenea, tiparele de activitate neuronală a populației. Între ele, celulele piramidale și celulele granulare sunt foarte liniștite (dar nu silențioase). În timpul unui val ascuțit, până la 5-10% din populație poate emite potențiale de acțiune pe parcursul unei perioade de 50 msec; multe dintre aceste celule nu emit unul, ci o rafală de vârfuri.
Aceste două moduri de activitate hipocapampală pot fi observate atât la primate, cât și la șobolani, cu excepția importantă că a fost dificil de observat o ritmicitate theta robustă în hipocampul primatelor. Cu toate acestea, există valuri ascuțite calitativ similare și schimbări similare dependente de stare în activitatea populației neuronale.
Ritmul theta
Din cauza straturilor sale neuronale dens împachetate, hipocampul generează unele dintre cele mai mari semnale EEG dintre toate structurile cerebrale. În unele situații, EEG-ul este dominat de unde regulate, care continuă adesea timp de mai multe secunde. Acest model EEG este cunoscut sub numele de ritm theta. Acesta a fost unul dintre primele fenomene EEG descoperite: prima descriere a fost făcută de Jung și Kornmuller, în 1938. Cu toate acestea, abia în 1954, odată cu publicarea de către Green și Arduini a unui studiu îndelungat și amănunțit al ritmului theta la iepuri, pisici și maimuțe, interesul a luat cu adevărat amploare.Poate în mare parte pentru că au legat ritmul theta de excitare, care era subiectul fierbinte al zilei, articolul lor a provocat o avalanșă de studii ulterioare, ceea ce a dus la publicarea a sute de studii despre fiziologia și farmacologia theta în anii 1950 și 1960. În ciuda acestui volum de muncă destul de impresionant, multe întrebări au rămas fără răspuns, în special cea a funcției. Chiar și în prezent, această întrebare, cea mai critică dintre întrebări, nu a primit încă un răspuns convingător.
Ritmicitatea Theta este foarte evidentă la iepuri și rozătoare și, de asemenea, este clar prezentă la pisici și câini. Dacă theta poate fi observată la primate este o întrebare supărătoare. Green și Arduini au raportat doar explozii foarte scurte de ritmicitate mai degrabă neregulată la maimuțe, iar majoritatea studiilor ulterioare au observat puțin mai mult. Cu toate acestea, variațiile de metodologie au făcut dificilă tragerea unor concluzii puternice.
La șobolani (animalele care au fost de departe cele mai mult studiate), theta este observată în principal în două condiții: în primul rând, atunci când un animal merge sau interacționează în alt mod activ cu împrejurimile sale; în al doilea rând, în timpul somnului REM. Frecvența crește în funcție de viteza de alergare, începând de la aproximativ 6,5 Hz în partea inferioară și crescând până la aproximativ 9 Hz în partea superioară, deși frecvențe mai mari sunt uneori observate în cazul unor mișcări dramatice, cum ar fi săriturile peste distanțe mari. La alte specii de animale, mai mari, frecvențele theta sunt în general puțin mai mici. Dependența comportamentală pare, de asemenea, să varieze în funcție de specie: la pisici și iepuri, theta este adesea observată în timpul stărilor de vigilență nemișcată. Acest lucru a fost raportat și în cazul șobolanilor, dar numai atunci când aceștia sunt foarte speriați.
Theta nu este limitată doar la hipocampus. La șobolani, ea poate fiobservată în multe părți ale creierului, inclusiv în aproape toate cele careinteracționează puternic cu hipocampul. Se crede că stimulatorul ritmului se află în zona septală medială: această zonă se proiectează în toate regiunile care prezintă ritmicitate theta, iar distrugerea ei elimină theta în tot creierul. (Ar putea exista o singură excepție, o mică zonă din hipotalamus numită supramamillarnucleus, care pare a fi capabilă să susțină theta independent de septum în anumite situații.)
Funcția theta, presupunând că are una, nu a fost încă explicată în mod convingător, deși au fost propuse numeroase teorii.Cea mai populară tendință a fost aceea de a o corela cu învățarea și memoria.Este bine stabilit faptul că leziunile septului medial–nodul central al sistemului theta–provoacă perturbări grave ale memoriei.Cu toate acestea, septul mediu este mai mult decât un simplu controlor al theta,este, de asemenea, principala sursă de proiecții colinergice cătrehipocampus. Nu s-a stabilit că leziunile septalei își exercită efectele în mod specific prin eliminarea theta.
Undele ascuțite
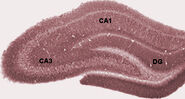
În timpul somnului sau în timpul stărilor de veghe, când un animal se odihnește sau în alt mod nu este implicat cu mediul înconjurător, EEG-ul hipocampal prezintă un model de unde lente neregulate, ceva mai mari în amplitudine decât undele theta. Acest model este ocazional întrerupt de valuri mari numite unde ascuțite. Aceste evenimente sunt asociate cu explozii de activitate de vârfuri, care durează 50-100 msec, în celulele piramidale din CA3 și CA1. Ele sunt, de asemenea, asociate cu oscilații EEG de înaltă frecvență de scurtă durată, numite “ondulații”. Ondulațiile, cu frecvențe în intervalul 150-200 Hz la șobolani, pot fi detectate, de obicei, numai de către electrozi localizați fie în interiorul, fie foarte aproape de stratul corpului celular CA1. În contrast, electrozii localizați oriunde în interiorul hipocampusului, sau chiar în structurile cerebrale învecinate, vor capta adesea undele ascuțite ca devieri EEG mari și lente, cu o durată de 200-400 msec.
La șobolani, undele ascuțite sunt cele mai robuste în timpul somnului, când apar la o rată medie de aproximativ 1 pe secundă, dar într-un tipar temporal foarte neregulat. Undele ascuțite apar, de asemenea, în timpul stărilor inactive de veghe, dar sunt mai puțin frecvente atunci și de obicei mai mici. Undele ascuțite au fost, de asemenea, observate în lobul temporal uman și în hipocampul maimuțelor.La maimuțe, undele ascuțite sunt destul de robuste, dar nu apar aproape la fel de frecvent ca la șobolani.
Unul dintre cele mai interesante aspecte ale undelor ascuțite este că ele par să fie asociate cu memoria. Wilson și McNaughton 1994, șinumeroase studii ulterioare, au raportat că atunci când celulele locului hipocampale au câmpuri de ardere spațiale care se suprapun (și, prin urmare, trag adesea în aproape simultaneitate), acestea tind să prezinte o activitate corelată în timpul somnului care urmează sesiunii comportamentale. S-a constatat că această intensificare a corelației, cunoscută în mod obișnuit sub numele de reactivare, este limitată în principal la undele ascuțite. S-a propus că undele ascuțite sunt, de fapt, reactivări ale modelelor de activitate neuronală care au fost memorate în timpul comportamentului, determinate de întărirea conexiunilor sinaptice în cadrul hipocampusului. Această idee formează o componentă cheie a teoriei “memoriei în două etape”, susținută de Buzsaki și alții, care propune că amintirile sunt stocate în hipocampus în timpul comportamentului și apoi transferate mai târziu în neocortex în timpul somnului: se sugerează că undele ascuțite determină schimbări sinaptice Hebbian în țintele neocorticale ale căilor de ieșire hipocampale.
Evoluție
Hipocampul are un aspect în general similar în toată gama de specii de mamifere, de la cele bazale, cum ar fi ariciul, la cele mai “avansate”, cum ar fi oamenii.Raportul dintre dimensiunea hipocampului și dimensiunea corpului crește în general, fiind de aproximativ două ori mai mare la primate decât la arici. Cu toate acestea, nu crește nici pe departe la o rată apropiată de cea a raportului neocortexului la dimensiunea corpului.Astfel, hipocampul ocupă un volum mult mai mare din cortex la rozătoare decât la primate.
Există, de asemenea, o relație generală între dimensiunea hipocampusului și memoria spațială: atunci când se fac comparații între specii similare, cele care au o capacitate mai mare de memorie spațială tind să aibă volume hipocampale mai mari…. Această relație se extinde, de asemenea, la diferențele de sex: în cazul speciilor în care masculii și femelele prezintă diferențe puternice în ceea ce privește capacitatea de memorare spațială, acestea tind, de asemenea, să prezinte diferențe corespunzătoare în ceea ce privește volumul hipocampului
Speciile nemamiferelor nu au o structură cerebrală care să semene cu hipocampul mamiferelor, dar au una care este considerată omoloagă. Hipocampul, așa cum s-a arătat mai sus,este în esență marginea mediană a cortexului. Numai mamiferele au cortexul complet dezvoltat, dar structura din care a evoluat, numită pallium, este prezentă la toate vertebratele, chiar și la cele mai primitive, cum ar fi lampreiul sau peștele hagiu. Palliul este de obicei împărțit în treizone: medială, laterală și dorsală. Palliul medial formează premergătorul hipocampusului. Acesta nu se aseamănă vizual cu hipocampul, deoarece straturile nu sunt deformate în formă de S și nici nu sunt învelite de girusul dentat, dar omologia este indicată de afinitățile chimice și funcționale puternice. Există acum dovezi că aceste stucturi asemănătoare cu cele ale hipocampului sunt implicate în cunoașterea spațială la păsări, reptile și pești.
La păsări, corespondența este suficient de bine stabilită pentru ca majoritatea anatomiștilor să se refere la zona paliativă medială ca fiind “hipocampul aviar”. Numeroase specii de păsări au abilități spațiale puternice,în special cele care ascund hrana. Există dovezi care arată că păsările care ascund hrană au un hipocampus mai mare decât alte tipuri de păsări și că afectarea hipocampusului cauzează deteriorarea memoriei spațiale….
Povestea peștilor este mai complexă. La peștii teleost (care alcătuiesc marea majoritate a speciilor existente), creierul anterior este ciudatdistorsionat în comparație cu alte tipuri de vertebrate. Cei mai mulți neuroanatomiști cred că creierul anterior al peștilor teleostești este în esență inversat, ca o șosetă întoarsă pe dos, astfel încât structurile care se află în interior, lângă ventricule, la majoritatea vertebratelor, se găsesc în exterior la peștii teleostești și viceversa. Una dintre consecințele acestui fapt este că se crede că palliul medial (zona “hipocampală”) al unei vertebrate tipice corespunde palliului lateral al unui pește tipic. S-a demonstrat experimental că mai multe tipuri de pești (în special peștii galbeni) au capacități puternice de memorare spațială, formând chiar “hărți cognitive” ale zonelor pe care le locuiesc. Există dovezi că o afectare a palliului lateral afectează memoria spațială (navigația pe distanțe lungi, cum ar fi orientarea către casă a somonilor, pare să se bazeze totuși pe mecanisme diferite).
Astfel, rolul regiunii hipocampale în navigație pare să fi început cu mult timp în urmă în evoluția vertebratelor, precedând diviziunile care au avut loc cu sute de milioane de ani în urmă. Nu se știe încă dacă palierul medial joacăun rol similar la vertebrate și mai primitive, cum ar fi rechinii și racilele, sau chiar la lamprei și peștișori. Unele tipuri de insecte și moluște, cum ar fi caracatița, au, de asemenea, capacități puternice de învățare spațială și de navigare, dar acestea par să funcționeze diferit de sistemul spațial al mamiferelor, astfel încât nu există încă niciun motiv întemeiat pentru a crede că acestea au o origine evolutivă comună; de asemenea, nu există o asemănare suficientă în structura creierului pentru a permite identificarea a ceva asemănător cu un “hipocampus” la aceste specii.
Vezi și
- Amigdalohippocampectomie
- Fășura mediană a creierului anterior
- Septal nuclei
Referințe & Bibliografie
Texte cheie
Cărți
- Aboitiz, F, Morales D, Montiel J (2003). Originea evolutivă a izocortexului mamiferelor: Towards an integrated developmental and functional approach. Behav. Brain Sciences 26: 535-552.
- Amaral, D; Lavenex P (2006). “Cap. 3. Hippocampal Neuroanatomy” Andersen P, Morris R, Amaral D, Bliss T, O’Keefe J The Hippocampus Book, Oxford University Press.
- Broglio, C, Gómez A, Durán E, Ocaña FM, Jiménez-Moya F, Rodríguez F, Salas C (2002). Semnele distinctive ale unui plan comun al vertebratelor vertebrate forebrain: Zonele paliene specializate pentru memoria spațială, temporală și emoțională la peștii actinopterygiani. Brain Res. Bull. 57: 397-399.
- Buzsáki, G (2002). Oscilațiile Theta în hipocampus. Neuron 33: 325-340.
- Buzsáki, G (2006). Ritmurile creierului.
- Cantero, JL, Atienza M, Stickgold R, Kahana MJ, Madsen JR, Kocsis B (2003). Oscilații theta dependente de somn în hipocampul și neocortexul uman. J. Neurosci. 23: 10897-10903.
- Colombo, M, Broadbent N (2000). Este hipocampul aviar un omolog funcțional al hipocampusului mamiferelor?”. Neurosci. Biobehav. Rev. 24: 465-484.
- Eichnbaum, H; Cohen NJ (1993). Memory, Amnesia, and the Hippocampal System, MIT Press.
- Ekstrom, AD, Kahana MJ, Caplan JB, Fields TA, Isham EA, Newman EL, Fried I (2003). Rețele celulare care stau la baza navigației spațiale umane. Nature 425: 184-188.
- Gray, JA; McNaughton N (2000). Neuropsihologia anxietății: An Enquiry into the Functions of the Septo-Hippocampal System, Oxford University Press.
- Green, JD, Arduini AA (1954). Activitatea electrică hipocampală în excitație. J. Neurophysiol. 17: 533-557.
- Jacobs, LF, Gaulin SJ, Sherry DF, Hoffman GE (1990). Evoluția cunoașterii spațiale: modelele de comportament spațial specifice sexului prezic dimensiunea hipocampului. PNAS 87: 6349-6352.
- Jacobs, LF (2003). Evoluția hărții cognitive. Brain Behav. Evol. 62: 128-139.
- Kirk, IJ, McNaughton N (1991). Focalizarea celulelor supramamilare și activitatea ritmică lentă hipocampală. Neuroreport 11: 723-725.
- Maguire, EA, Gadian DG, Johnsrude IS, Good CD, Ashburner J, Frackowiak RS, Frith CD (2000). Schimbări structurale legate de navigare în hipocampul șoferilor de taxi. PNAS 97: 4398-4403.
- McNaughton, BL, Battaglia FP, Jensen O, Moser EI, Moser MB (2006). Integrarea căilor și baza neuronală a “hărții cognitive”. Nat. Rev. Neurosci. 7: 663-678.
- Moser, EI, Kropf E, Moser M-B (2008). Place Cells, Grid Cells, and the Brain’s Spatial Representation System. Ann. Rev. Neurosci. 31.
- Nieuwenhuys, R (1982). An Overview of the Organization of the Brain of Actinopterygian Fishes. Am. Zool. 22: 287-310.
- O’Kane, G, Kensinger EA, Corkin S (2004). Dovezi pentru învățarea semantică în amnezia profundă: O investigație cu pacientul H.M.. Hippocampus 14: 417-425.
- O’Keefe, J; Nadel L (1978). The Hippocampus as a Cognitive Map, Oxford University Press.
- Portavella, M, Vargas JP, Torres B, Salas C (2002). Efectele leziunilor palice telencefalice asupra învățării spațiale, temporale și emoționale la peștii aurii. Brain Res. Bull. 57: 397-399.
- Rodríguez, F, Lópeza JC, Vargasa JP, Broglioa C, Gómeza Y, Salas C (2002). Memoria spațială și palierul hipocampal prin evoluția vertebratelor: perspective de la reptile și pești teleost. Brain Res. Bull. 57: 499-503.
- Sainsbury, RS, Heynen A, Montoya CP (1987). Corelații comportamentale ale hipocampului de tip 2 theta la șobolan. Fiziologie și comportament. 39: 513-519.
- Scoville, WB, Milner B (1957). Pierderea memoriei recente după leziuni hipocampale bilaterale. J. Neurol. Neurosurg. Psych. 20: 11-21.
- Shettleworth, SJ (2003). Memoria și specializarea hipocampală la păsările care stochează alimente: Challenges for Research on Comparative Cognition (Provocări pentru cercetarea cogniției comparative). Brain Behav. Evol. 62: 108-116.
- Skaggs, WE, McNaughton BL, Wilson MA, Barnes CA (1996). Precesia fazei Theta în populațiile neuronale hipocampale și comprimarea secvențelor temporale. Hippocampus 6: 149-176.
- Squire, LR (1992). Memoria și hipocampul: o sinteză din descoperirile făcute la șobolani, maimuțe și oameni. Psych. Rev. 99: 195-231.
- Squire, LR; Schacter DL (2002). The Neuropsychology of Memory (Neuropsihologia memoriei), Guilford Press.
- Vanderwolf, CH (1969). Activitatea electrică hipocampală și mișcarea voluntară la șobolan. EEG & Clin. Neurophysiol. 26: 407-418.
- Vargas, JP, Bingman VP, Portavella M, López JC (2006). Telencefalul și spațiul geometric la peștele auriu. Eur. J. Neurosci. 24: 2870-2878.
- West, MJ (1990). Stereological studies of the hippocampus: a comparison of the hippocampal subdivisions of diverse specii, inclusiv arici, rozătoare de laborator, șoareci sălbatici și oameni…. Prog. Brain Res. 83: 13-36.
Papers
- Amaral DG și Cowan WM. 1980. Aferențe subcorticale la formațiunea hipocampală la maimuță. Jurnalul de neurologie comparată. Feb 15; 189(4):573-91.
- Duvernoy, H. (2005) The Human Hippocampus, 3rd ed. Berlin: Springer-Verlag.
Material adițional
Lucrări
- Google Scholar
- http://www.psycheducation.org/emotion/hippocampus.htm Tururi minunate ale creierului!!!
- BrainMaps at UCDavis hippocampus
- Artificial Hippocampus
- BrainInfo at the University of Washington hier-164
- John O’Keefe & Lynn Nadel (1978) The Hippocampus as a Cognitive Map , Oxford University Press. Full Text
Imagini suplimentare
Adaugați o fotografie în această galerie
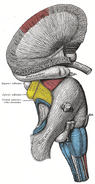
Creierul uman: Sistemul limbic
Amigdala – Girusul cingular – Girusul forțat – Hipocampul – Hipotalamusul – Corpul mamilar – Nucleul accumbens – Cortexul orbitofrontal – Girusul parahipocampal
|
v-d-e
Creierul uman: creierul anterior (trunchiul cerebral – cortexul cerebral – emisferele cerebrale, substanța cenușie) (TA A14.1.09.002-240, 301-320, GA 9.818-826) |
|||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lobul frontal |
|
||||||||||||||||
| Lobul parietal |
|
Superolateral
|
Lobulul mediu/inferior
|
Ambele
|
|||||||||||||
| Lobul occipital |
|
Superolateral
|
Medial/inferior
|
||||||||||||||
| Lobul temporal |
|
Superolateral
|
Girusul median/inferior
|
||||||||||||||
| Sulci/fisuri interlobare |
. |
Superolateral
|
Medială/inferioară
|
||||||||||||||
| Lobul limbic |
|
Alte
|
|||||||||||||||
| Lobul insular |
Girusul lung al insulei – Girusul scurt al insulei – Sulcusul circular al insulei |
||||||||||||||||
| General |
Operculum – Polii emisferelor cerebrale |
||||||||||||||||
Unele clasificări sunt aproximări, iar unele arii Brodmann se întind pe gyri.
|
|||||||||||||||||
Această pagină folosește conținut licențiat Creative Commons de pe Wikipedia (vezi autori).