- Introduction
- Metoder
- Distributionsdata og bioklassifikation
- Sammensætning af arter blandt biomer
- Tegenskabsdata
- Måling af egenskabshypervolumer
- Funktionel særprægethed og udbredelse
- Funktionelt rum og biome lighed
- Resultater
- Diskussion
- Knowledge Shortfalls
- Fælles mønstre inden for biomer
- Sammenligninger mellem biomer
- Konklusioner
- Datatilgængelighed
- Author Contributions
- Funding
- Interessekonflikterklæring
- Akkommentarer
- Supplementært materiale
Introduction
Ekologer og biogeografer inddeler biodiversiteten af landplanter i klimatisk bestemte biomes, med fysiognomier karakteriseret ved de dominerende arters vækstformer og funktionelle træk (Moncrieff et al., 2016). Biombegrebet har faktisk en lang historie, der går tilbage til Carl Ludwig Willdenow og Alexander von Humboldt. Willdenow erkendte, at lignende klimaer understøtter lignende vegetationsformer, og Humboldt observerede den udbredte sammenhæng mellem plantefordeling, fysiognomi og miljøfaktorer. Overordnet set afspejler biombegrebet den antagelse, at ensartede miljøbelastninger vælger arter med ensartede funktionelle egenskaber, uafhængigt af deres evolutionære historie. Samtidig er Jordens eksisterende biomes til en vis grad fylogenetisk forskellige, idet mange/flertallet af de karakteristiske arter stammer fra specifikke slægter, der udviser vigtige tilpasninger ikke blot til klimatisk selektion, men også til yderligere pres som f.eks. brand eller megaherbivorer (Woodward et al., 2004; Pennington et al., 2006; Donoghue og Edwards, 2014). Fordi biomerne repræsenterer regelmæssigheder i bred skala i Jordens vegetation, er forståelsen af funktionelle forskelle mellem biomerne af afgørende betydning for modellering af den globale kulstofcyklus og jordsystemets funktion, herunder reaktioner på antropogene globale ændringer (Bonan et al, 2012; van Bodegom et al., 2014; Xia et al., 2015).
I de seneste to årtier er der sket en hurtig vækst i forståelsen af den funktionelle dimension af plantebiodiversitet på tværs af kontinentale skalaer (Swenson et al., 2012; Lamanna et al., 2014; Šímová et al., 2018). Omfattende datasyntese- og analyseindsatser har defineret et begrænset funktionelt egenskabsrum, der inkorporerer variation i den samlede plantestørrelse, bladøkonomi og hydraulik, kendt som funktionelle egenskabsspektrer (Wright et al., 2004; Díaz et al., 2016). Funktionelle egenskabsspektrer definerer fysiologiske og økologiske afvejninger, der bestemmer planternes livshistoriske strategier (Westoby, 1998; Reich et al., 2003; Craine, 2009) og deres indflydelse på samfundssammensætning (Kraft et al., 2008), økosystemfunktion (Lavorel og Garnier, 2002; Garnier et al., 2004; Kerkhoff et al., 2005; Cornwell et al., 2008) og endda udviklingshastigheder (Smith og Donoghue, 2008; Smith og Beaulieu, 2009). Genereliteten og anvendeligheden af plantefunktionelle egenskabsspektrer har fremskyndet deres indarbejdelse i modeller for den globale fordeling af vegetation (van Bodegom et al., 2014), biogeokemisk cyklus (Bonan et al., 2012) og økosystemtjenester (Díaz et al., 2007; Cadotte et al., 2011; Lavorel et al., 2011). Der er dog kun få undersøgelser, der har undersøgt omfattende målinger af funktionel diversitet på biomskalaen på tværs af både dominerende og underordnede arter og livsformer.
Der er to modsatrettede sæt af forudsigelser om, hvordan funktionel diversitet varierer mellem klimatisk og fysiognomisk forskellige biomes. På den ene side, hvis biomers biodiversitet afspejler variationen i det tilgængelige økologiske nicherum, bør mindre taksonomisk forskelligartede biomer repræsentere en mindre, stort set indlejret delmængde af det funktionelle rum, der er besat af mere forskelligartede biomer. På den anden side kan sammensætninger, der besætter forskellige miljøer, på grund af den globale karakter af grundlæggende afvejninger i planters struktur og funktion og lignende selektionspres, der virker på samme måde på tværs af arter, faktisk dele lignende områder af egenskabsrummet (Reich et al., 2003; Wright et al., 2004; Díaz et al., 2016). Nyere undersøgelser bekræfter, at forholdet mellem klima og funktionel og taksonomisk diversitet (dvs. artsrigdom) er komplekst og skalaafhængigt. I nogle tilfælde følger den funktionelle diversitet nøje de klimatiske gradienter med lavere funktionel diversitet under mere varierende og ekstreme forhold (Swenson et al., 2012; de la Riva et al., 2018), og omfanget af funktionelt rum i lokale sammensætninger udvides med stigende taksonomisk rigdom (Lamanna et al., 2014; Li et al., 2018). Analyser af artspuljer på regional skala tyder dog på, at store ændringer i artsrigdommen kan være forbundet med minimale virkninger på den funktionelle diversitet (Lamanna et al., 2014; Šímová et al., 2015). Desuden kan den taksonomiske og funktionelle diversitets reaktion på klimaet være forskellig blandt større plantevækstformer (Šímová et al., 2018), og den relative diversitet af forskellige vækstformer kan ændre sig væsentligt langs de klimagradienter, der definerer forskellige biomer (Engemann et al., 2016).
Fremskridt i analysen af funktionel diversitet på kontinental til global skala er blevet begrænset til dels af datamangler i katalogiseringen af de taksonomiske, distributionelle, fylogenetiske og funktionelle aspekter af biodiversitet (Hortal et al., 2015). Desuden er der, selv i betragtning af de begrænsede tilgængelige data, betydelige informatiske udfordringer forbundet med at standardisere og integrere store, uensartede datasæt, der beskriver arternes geografiske fordeling, funktionelle træk og fylogenetiske relationer (Violle et al., 2014). Her undersøger vi fordelingen af funktionel diversitet af plantearter på tværs af biomerne i Nord- og Sydamerika ved hjælp af databasen Botanical Information and Ecology Network (BIEN; Enquist et al., 2016; Maitner et al., 2018), som samler oplysninger om fordeling og funktionelle træk på >100,000 arter af landplanter. Specifikt undersøger vi variationen i den funktionelle diversitet og distinktivitet af biomes, baseret på landplantearternes fordelingskort på en skala på 100 × 100 km gitterceller og et omfattende datasæt af seks funktionelle egenskaber, der afspejler de vigtigste akser af variation i økologiske strategier.
Vores mål i denne undersøgelse er 3-foldige. For det første dokumenterer vi for at fremhæve vedvarende datamangler omfanget af de tilgængelige data, der karakteriserer biomers funktionelle mangfoldighed og særpræg. For det andet karakteriserer vi på baggrund af de tilgængelige data fordelingen af den funktionelle mangfoldighed inden for biomerne på tværs af både dominerende og underordnede vækstformer. Disse analyser giver os mulighed for bedre at kvantificere et biomes funktionelle særpræg ved at identificere de mest almindelige funktionelle strategier for de mest udbredte arter inden for det. For det tredje spørger vi, om biomerne faktisk er karakteriseret ved funktionelt forskellige samlinger af arter, baseret på overlapningen af flerdimensionale hypervolumer i funktionelle egenskabsrum.
Metoder
Distributionsdata og bioklassifikation
For at reducere virkningerne af prøvetagningsbias, der er karakteristisk for datasæt for forekomst, der er samlet fra flere ressourcer, brugte vi BIEN 2.0-områdekortene for 88.417 af plantearter fordelt i Nord- og Sydamerika (Goldsmith et al, 2016). BIEN-databasen integrerer standardiserede planteobservationer, der stammer fra herbariums eksemplarer og opgørelser af vegetationsområder. Artsudbredelseskort i BIEN-databaserne blev estimeret ved hjælp af en af tre fremgangsmåder, afhængigt af de forekomster, der er tilgængelige for hver art. For arter med kun en eller to forekomster (ca. 35 % af arterne) blev det geografiske udbredelsesområde defineret som et kvadratområde på 75 000 km2 omkring hvert datapunkt. De geografiske områder for arter med tre eller fire forekomster blev identificeret ved hjælp af et konvekse skrog (ca. 15 % af arterne). Endelig blev udbredelseskortene for arter med mindst fem forekomster fremstillet ved hjælp af Maxent-algoritmen til modellering af artsfordeling, hvor 19 klimalag blev anvendt som forudsigelsesvariabler og 19 rumlige egenvektorer som filtre til at begrænse modellernes forudsigelser (se Goldsmith et al, 2016 for yderligere oplysninger om metodologi for udbredelseskort).
Vi overlejrede BIEN 2.0-plantearternes kort på et 100 × 100 km gitterkort med en Lambert Azimuthal Equal Area-projektion for at opnå en tilstedeværelses-/fraværsmatrix for arter for hver gittercelle. På grundlag af Olson et al. (2001) biome-klassificeringen tildelte vi hver matrixgittercelle til en af biomkategorierne. På grund af beregningsmæssige begrænsninger i forbindelse med de efterfølgende analyser slog vi nogle biomes sammen på grundlag af deres ligheder med hensyn til klima og vegetation og litteratur for at opnå en bred klassifikation som beskrevet i tabel 1 (undtagen indlandsvand, klippe og is og mangrove). Chaco- og Caatinga-økoregionerne blev klassificeret som henholdsvis Xeric Woodlands og Dry forest i henhold til Prado og Gibbs (1993), Pennington et al. (2000), Banda et al. (2016), Silva de Miranda et al. (2018).
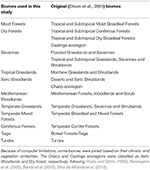
Tabel 1. Oversigt over den bioklassifikation, der er vedtaget i denne undersøgelse, og den tilsvarende (Olson et al., 2001) bioklassifikation (eksklusive indlandsvand, klippe og is og mangrove).
Sammensætning af arter blandt biomer
For at forstå variationen i funktionelle egenskabsrum blandt biomerne undersøgte vi først forskellene i plantearternes sammensætning blandt dem. På grundlag af artslisten for hver gittercelle og hvert biome definerede vi hvert bioms karakteristiske arter som de arter, der har den maksimale andel af deres udbredelsesområde i det pågældende biome. Disse lister over geografisk fremherskende arter i et givet biome blev sammenlignet med listen over arter i andre biomes. Denne parvise sammenligning giver en enkel måde at udlede en retningsbestemt taksonomisk overlapning mellem biomerne (dvs. andelen af fremherskende arter i et biom, der deles med et andet biom). Af de 88.417 arter med tilgængelige udbredelseskort har 44.899 arter udbredelsesområder, der spænder over mere end ét biome, og 43.518 arter er endemiske for et specifikt biome.
Tegenskabsdata
Vi ekstraherede alle egenskabsoplysninger for plantearter i den nye verden, der er tilgængelige i BIEN 3.0-datasættet (hentet den 7. februar 2018), hvilket giver i alt 80.405 observationer af egenskaber på artsniveau. Vi filtrerede derefter oplysningerne for seks funktionelle egenskaber: maksimal plantehøjde (m), frømasse (mg), trætæthed (mg/cm3), specifikt bladareal SLA (cm2/g) og bladfosfor- og bladkvælstofkoncentration pr. masseenhed (mg/g). Efter filtrering var der i alt 18 192 observationer på artsniveau tilbage. Heraf blev den delmængde på 8 820 arter med både udbredelseskort og egenskabsoplysninger anvendt til yderligere analyser.
For at estimere det funktionelle egenskabsrum for hvert biome krævede vi fuldstændige egenskabsdata. Af denne grund imputerede vi fylogenetisk manglende egenskabsdata (Bruggeman et al., 2009; Penone et al., 2014; Swenson, 2014; Swenson et al., 2017) ved hjælp af R-pakken “Rphylopars” v 0.2.9 (Goolsby et al., 2017) og den nyligt offentliggjorte fylogeni af frøplanter af Smith og Brown (2018) som en basislinje. Fylogenetisk imputering er et værktøj til at forudsige manglende data i datasæt med funktionelle egenskaber baseret på den antagelse, at nært beslægtede arter har tendens til at have lignende egenskabsværdier (Swenson, 2014). Vi brugte ALLBM-træet (dvs. genbank og Open Tree of life taxa med en rygsøjle leveret af Magallón et al., 2015), fordi det maksimerede overlapningen mellem arter med tilgængelige egenskabs- og fordelingsoplysninger. Efter egenskabsimputationen var der i alt 7.842 arter med komplette egenskabsoplysninger og udbredelseskort tilbage til yderligere analyse.
Måling af egenskabshypervolumer
Vi målte den relative funktionelle diversitet af biomer ved at beregne egenskabshypervolumer fra artspuljer inden for biomceller. På grund af beregningsmæssige begrænsninger konstruerede vi egenskabshypervolumerne ved hjælp af en tilfældig stikprøve på 20 % af cellerne i hvert biome. Hypervolumerne for hver gittercelle blev estimeret ved hjælp af de ekstraherede seks funktionelle træk og R-pakken “hypervolume” (Blonder et al., 2014, 2018) ved hjælp af den Gaussiske KDE-metode med standard Silverman-båndbreddeestimator. Frømasse, højde og trætæthed blev log-transformeret, mens SLA blev kvadratrodstransformeret. Alle egenskaber blev skaleret og centreret før analysen. Hypervolumer rapporteres i enheder af standardafvigelser (sd) til potensen til antallet af anvendte egenskaber (dvs. sd6).
Funktionel særprægethed og udbredelse
Da biomerne er karakteriseret ved deres dominerende vegetation, undersøgte vi også den geografiske udbredelse og funktionelle særprægethed af arter inden for biomerne. Ved hjælp af kort over arternes udbredelsesområde og oplysninger om funktionelle egenskaber estimerede vi den funktionelle særprægethed og udbredelse af hver art i hvert biome i henhold til den konceptuelle ramme for funktionel sjældenhed af Violle et al. (2017). Ved hjælp af hele sættet af (målte og imputerede) egenskaber målte vi først den euklidiske afstand i det standardiserede egenskabsrum mellem alle par af arter. Derefter beregnede vi den funktionelle distinktivitet (Di) for hver art i hvert biome som den gennemsnitlige funktionelle afstand mellem en art og de N andre arter inden for biomeartspuljen. Di blev skaleret mellem 0 og 1 inden for hvert biome, idet lavere værdier repræsenterer arter, der er funktionelt fælles (redundante), og højere værdier repræsenterer arter, der er funktionelt særprægede i forhold til de andre arter i biomet. Vi estimerer også den geografiske udbredelse (Wi) af en art i et biome, målt som antallet af gitterceller, der er besat af arten i et biome, i forhold til det samlede antal celler i det pågældende biome. En værdi på 1 angiver, at den pågældende art er til stede i alle de gitterceller, der er dækket af biomet. Både funktionel særpræg og geografisk udbredelse blev beregnet ved hjælp af R-pakken “funrar” (Grenié et al., 2017).
Da vi mangler omfattende mål for dominans baseret på lokal forekomst eller biomasse, brugte vi målene Di og Wi til for hvert biome at identificere de mest “almindelige” arter som dem, der både er geografisk udbredte (Wi > 0,5) og funktionelt ensartede (Di < 0,25). Denne sidste tærskel blev anvendt, da den tredje kvantil af værdierne for funktionel særprægethed blandt biomerne varierede fra 0,2 til 0,3. For at skelne funktionelt forskellige arter fra funktionelt ensartede arter i de efterfølgende analyser anvendte vi derfor en værdi på 0,25 som grænseværdi (dvs, arter med distinktivitetsværdier < 0,25 blev betragtet som funktionelt fælles eller redundante inden for et specifikt biome).
Funktionelt rum og biome lighed
For at estimere overlapningen mellem biomers hypervolumer anvendte vi Sørensen lighedsindekset ved hjælp af (i) det samlede antal arter og (ii) listen over arter, der betragtes som funktionelt fælles og geografisk udbredte for hvert biome. Ligheden mellem funktionelle træk blev beregnet som det parvise fraktionelle overlap af hypervolumer mellem biomerne. Den fraktionelle overlapning blev beregnet ved at dividere det dobbelte af volumenet af skæringspunktet mellem to hypervolumer med volumenet af deres unioner. Alle hypervolumer blev estimeret ved hjælp af R-pakken “hypervolume” (Blonder et al., 2014, 2018) ved hjælp af den Gaussiske KDE-metode med standard Silverman-båndbreddestimator.
Resultater
Vi fandt et betydeligt overlap i sammensætningen af plantearter på tværs af biomerne, baseret på skæringspunktet mellem de modellerede artsområder (se figur 1 og supplerende tabel 1). Denne taksonomiske overlapning er større inden for tropiske og tempererede biomer, med relativt få arter, der deles mellem disse to klimazoner. Det er interessant, at xeriske skovområder deler arter med både tropiske og tempererede biomer. På trods af den høje andel af arter, der er karakteristisk for tropisk fugtig skov (83 %), deler dette biome også et stort antal arter med andre tropiske biomes som f.eks. tørre skove, savanner, xerotiske skove og tropiske græsmarker. Den store taksonomiske overlapning af tempererede biomes såsom tempererede græsmarker, middelhavsskove, taiga og tundra tyder på, at der er mulighed for lav funktionel særprægethed, idet nogle tempererede biomes repræsenterer en fattigere delmængde af de mere artsrige, funktionelt mangfoldige tropiske biomes.
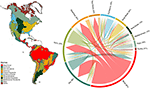
Figur 1. Overlapning af plantearter mellem biomerne i den nye verden. Procentværdierne udtrykker den andel af de arter, der forekommer i et biome, og som har den største del af deres geografiske udbredelsesområde i det pågældende biome, N = 84 413. Basen af hver forgrenet pil er placeret således, at den viser det biome, der omfatter den største andel af en arts udbredelsesområde, mens bredden repræsenterer det antal arter, der deles med de biomes, der ligger i pilens spidser. Se Supplerende tabel 1 for den underliggende matrix.
Med mindre end 10 % af de kortlagte arter repræsenteret var egenskabsdataene ret sparsomme for vores artssammensætninger på biometrisk skala (figur 2A). Desuden varierede de tilgængelige egenskabsdata betydeligt på tværs af egenskaber, planteklasser og biomer. Fylogenetisk set var blad-P og trætæthed særligt dårligt repræsenteret, idet hele klasser ikke havde nogen tilgængelige data. Som følge heraf måtte mange af de ca. 2/3 af egenskabsværdierne for vores 7.842 arter, som skulle imputeres, tildeles i mangel af nært beslægtede taxa. Geografisk set er SLA, højde og frømasse underprøvet i troperne og det tempererede Sydamerika, mens blad-N og trætæthed er mere jævnt fordelt i de forskellige regioner. Blad-P er dårligt opsamlet på tværs af alle biomer. Trait-hypervolumer skabt fra artspooler af et tilfældigt udvalg på 20 % af cellerne for hvert biome viser hovedsagelig forskelle mellem tropiske og tempererede/koldt tempererede biomer (figur 2B). Der er en tendens til større og mere varierende hypervolumer i tropiske biomer med den største variation i fugtige og tørre tropiske skove. Blandt de tempererede/kolde biomer er det nåletræer og tempererede blandede skove, der har de største hypervolumer af egenskaber. Det er interessant, at xeriske buskmarker udviser en fordeling af hypervolumer, der minder mere om tropiske end tempererede biomer.
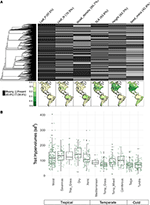
Figur 2. (A) Andel af arter i BIEN 3.0-databasen med kendte (grå) eller manglende (sorte) egenskabsværdier. Fylogeni til venstre svarer til frøplanternes ALLBM-træ fra Smith og Brown (2018) (dvs. GenBank og Open Tree of life taxa med en rygsøjle leveret af Magallón et al., 2015). Kort repræsenterer andelen af arter med egenskabsoplysninger i forhold til det samlede antal arter i BIEN 2.0-databasen. (B) Fordeling af egenskabshypervolumer af 20 % af tilfældigt udvalgte 100 × 100 km celler i hvert biome. Hypervolumer er rapporteret i enheder af standardafvigelser i potens af det anvendte antal træk.
Forholdet mellem funktionel særpræg og geografisk fordeling er bemærkelsesværdigt ens blandt biomerne (figur 3). I alle biomer er langt størstedelen af arterne både geografisk begrænsede og funktionelt ensartede. Samtidig var de mest funktionelt særprægede arter inden for hvert biome generelt også geografisk begrænsede, og de mest geografisk udbredte arter var næsten altid funktionelt ens.
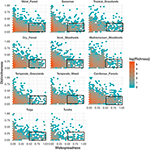
Figur 3. Mønstre af funktionel særprægethed på tværs af biomer. Distinktivitet repræsenterer, hvor langt arter er funktionelt fra hinanden inden for et biome (dvs. den gennemsnitlige parvise fænotypiske afstand fra en fokusart til alle de andre). Jo større værdien er, jo mere fjernt er en art fra centrum af biomets funktionelle område. Udbredelse måler, hvor geografisk udbredt en art er inden for et biome. En værdi på 0 angiver, at en art kun findes i en enkelt biocelle. Bin-værdierne er antallet af arter på en loge-skala. Rektanglerne repræsenterer de arter, der betragtes som funktionelt ensartede og udbredte i hvert biome ved hjælp af cut-off-værdier på henholdsvis 0,25 og 0,5 for distinktivitet og udbredelse.
Vi fandt betydelig variation i den proportionelle fordeling af vækstformer inden for og mellem biomerne. Træarter udgør de dominerende vækstformer i tropiske biomer med hensyn til antal arter, mens urteagtige arter dominerer i tempererede miljøer (figur 4). Når vi kun tog hensyn til de arter, der er udbredte og funktionelt almindelige i hvert biome, ændrede fordelingen af vækstformer på tværs af biomerne sig, især hvad angår andelen af træer, urter og græsser. I tropiske biomer faldt andelen af træer f.eks. i alle biomer undtagen i fugtig skov. I tempererede biomer steg andelen af græsser, især mod polarområderne. Interessant nok ligner fordelingen af vækstformer i xeriske skove i højere grad fordelingen i tempererede biomer, når der kun tages hensyn til udbredte og funktionelt almindelige arter.
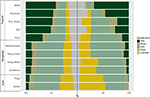
Figur 4. Fordeling af vækstformer i hvert biome (venstre) ved hjælp af det samlede antal arter og (højre) ved hjælp af kun de arter, der er funktionelt ensartede og udbredte inden for biomet (se figur 3).
Parvise sammenligninger af artssammensætningen mellem biomerne afslører tre hovedgrupper, der repræsenterer de tropiske, tempererede og polare klimazoner (figur 5A), hvilket afspejler den store taksonomiske overlapning inden for, men ikke mellem disse regioner (figur 1). Parvise sammenligninger af egenskabshypervolumer mellem biomerne viser en mindre klar klynge af klimazoner (figur 5B). I dette tilfælde overlapper det funktionelle rum for xeriske skovområder betydeligt med tempererede biomer. Denne analyse afslører også, at taigas funktionelle rum overlapper med tempererede græsmarker og blandede skove. Den parvise sammenligning af egenskabshypervolumener mellem biomerne ved kun at anvende de arter, der betragtes som funktionelt almindelige og udbredte, viser mindre overlapning i egenskabsrummene mellem og inden for klimazonerne (figur 5C). Men selv om xeriske skove nu er grupperet sammen med resten af de tropiske biomer, udviser disse habitater sammen med tropiske græsarealer et stort overlap i funktionelt rum med tempererede biomer såsom middelhavsskove og tempererede græsarealer.
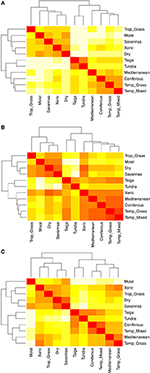
Figur 5. (A) Parvis ulighed i artssammensætningen mellem biomerne. (B) Parvis ulighed i egenskabshypervolumer (1-Sørensen-sammenfald) mellem biomer ved hjælp af det samlede antal arter. (C) Parvis ulighed i egenskabshypervolumer (1-Sørensen-sammenfald) mellem biomerne ved kun at anvende de arter, der anses for at være funktionelt ens og udbredte. Jo lysere celle, jo større er forskelligheden.
Diskussion
Vores analyser giver tre vigtige indsigter til forståelse af terrestriske biomes gennem en funktionel linse. For det første viser vi, at der på trods af fremskridt i indsamlingen og syntesen af primære biodiversitetsdata fortsat er betydelige vidensmangler, som kan begrænse vores evne til at kvantificere den funktionelle biodiversitet i biomerne på kontinental til global skala. For det andet viser vores analyser af de tilgængelige data ikke desto mindre, at alle biomerne i Nord- og Sydamerika har et bemærkelsesværdigt fælles mønster, hvor de geografisk mest udbredte arter i et biome har tendens til at dele fælles funktionelle træk, mens de mest funktionelt særprægede arter altid er begrænset i deres udbredelse. For det tredje kan man, når man kun tager hensyn til de udbredte og funktionelt almindelige arter, lettere skelne biomerne fra hinanden i funktionelt henseende, og mønstre af forskellighed mellem biomerne synes at afspejle en overensstemmelse mellem klima og planternes funktionelle nicherum. Samlet set tyder vores resultater på, at mens undersøgelsen af biomers funktionelle diversitet stadig er i sin indledende fase, vil en yderligere udvikling af feltet sandsynligvis give indsigt, der forbinder evolution, biogeografi, samfundssammensætning og økosystemfunktion.
Knowledge Shortfalls
BIEN-databasen er specifikt designet til at lukke huller i vores viden om plantebiodiversitet, men som Hortal et al. (2015) påpeger, fører interagerende videnmangler til usikkerhed ved kvantificering af biodiversitet på de største skalaer. For eksempel havde ikke alene mindre end 10% af vores kortlagte arter både egenskabsdata og fylogenetiske oplysninger til rådighed, men disse manglende egenskabsdata (~66% af arternes egenskabsværdier) var ret strukturerede, både fylogenetisk og geografisk. Således interagerer den Raunkiaeriske mangel på funktionelle egenskabsdata (sensu Hortal et al., 2015) med den Wallaceanske mangel på distributionsinformation og den Darwinistiske mangel på fylogenetisk forståelse. Og selv om vi kan bruge fylogenetisk viden til rimeligt at imputere disse manglende værdier (Swenson et al., 2017), forbliver flere aspekter af de resulterende mønstre, som er vigtige for bioklassificering, usikre. For eksempel tyder de eksisterende data om vækstformer på, at træagtige arter dominerer i alle tropiske biomer, mens den proportionelle diversitet af urteagtige vækstformer er meget højere i tempererede og polære biomer (se også Engemann et al., 2016; Zanne et al., 2018). Er denne breddegradsstigning i den relative diversitet af urteagtige planter udtryk for prøvetagningsbias og manglende taksonomisk viden om tropiske urteagtige planter, eller afspejler den de forskellige evolutionære og biogeografiske historier i de Nearktiske og Neotropiske riger? Dominerende vækstformer er et af de vigtigste kendetegn for biomer, så systematiske ændringer i de mest forskelligartede vækstformer (uanset om de er dominerende eller ej) vil nødvendigvis påvirke vores forudsigelser om, hvordan den funktionelle diversitet kan ændre sig på tværs af biomer, og hvordan disse ændringer vil påvirke økosystemets funktion og tjenester.
Vi brugte kort over arternes udbredelsesområde som inputdata primært for at overvinde det ubestridelige problem med prøvetagningsbias, der er karakteristisk for datasæt, der er indsamlet fra flere kilder (se supplerende figur 1). Udbredelseskort, der er udarbejdet ud fra modeller for artsfordeling, kan imidlertid udgøre et yderligere element af usikkerhed i vores resultater. I betragtning af den geografiske opløsning i denne undersøgelse og den rumlige kompleksitet i visse biomes kan disse modeller f.eks. have overvurderet den geografiske udbredelse af nogle arter fra celler i et enkelt biome til celler i nærliggende og indbyrdes forbundne biomer. Anvendelsen af data om forekomst ændrede ikke de generelle resultater af denne undersøgelse (se supplerende figurer 2-6); en mindre reduktion i det funktionelle overlap mellem biomerne kunne dog faktisk afsløre en overvurdering af nogle arters udbredelsesområder (se supplerende figurer 2, 5 og 6). Anvendelse af oplysninger om forekomst ændrede også de generelle forhold mellem funktionel særpræg og udbredelse i vores resultater på grund af den relative overudvælgelse af tempererede vs. tropiske regioner (se supplerende figur 4). I dette tilfælde er vores mål for udbredelse begrænset af den ekstremt begrænsede prøveudtagning af de fleste arter i tropiske biomer. Efterhånden som datasættene og prøveudtagningen forbedres, vil der sandsynligvis være mulighed for at reducere disse usikkerheder i fremtiden.
Den anden datamangel, der skal afhjælpes for at forstå biomers funktionelle diversitet, er den prestoniske mangel på oplysninger om forekomst. Abundans er særlig vigtig i undersøgelser af funktionel diversitet, fordi de træk, der har størst betydning for samfundssammensætningen, økosystemets funktion og den biogeokemiske cyklus, er de mest dominerende arters træk. Fordi vi baserede vores sammensætninger på rækkeviddekort afledt af artsfordelingsmodeller, kunne vi kun behandle den geografiske komponent af almindelighed, men det positive forhold mellem lokal forekomst og geografisk rækkevidde, den såkaldte belægnings-overflodssammenhæng, antyder, at de mest udbredte arter i hvert biome ofte også kan være blandt de mere rigelige (Borregaard og Rahbek, 2010; Novosolov et al., 2017). Da biomer generelt er karakteriseret ved de dominerende vækstformer i regionen, kan integrering af oplysninger om forekomst resultere i biomer, der viser mindre funktionel overlapning. Men selv om BIEN har indsamlet omfattende plottedata, der giver mulighed for at estimater af lokal abundans, er det vanskeligt at foretage rimelige sammenligninger på tværs af forskellige regioner og vækstformer på grund af ujævn prøvetagning, regionale forskelle i gammadiversitet og inkommensurable metoder til kvantificering af abundans.
Fælles mønstre inden for biomer
På trods af de vedvarende huller i de tilgængelige data afdækkede vores analyser et tidligere udokumenteret forhold, der er fælles for alle biomerne på den vestlige halvkugle: I ethvert biome har de mest udbredte arter også en tendens til at udvise lav funktionel særpræg, mens de mest funktionelt særprægede arter næsten altid er begrænset i deres udbredelse (figur 3). Dette forhold mellem “belægning og redundans” kan tyde på, at de klimatiske forhold, der er fremherskende i et biome, vælger et sæt fælles karakteristika, idet de mere funktionelt særprægede arter er begrænset til en sjældnere delmængde af habitater i biomet. Den generelle forekomst af funktionelt ensartede arter i alle biomer og “belægning-redundans”-forholdet er begge i overensstemmelse med en nyere global analyse af funktionel diversitet på samfundsniveau, der tyder på, at filtrering af levesteder fører til sameksistens af funktionelt ensartede arter (Li et al., 2018) samt undersøgelser, der viser, at funktionelle redundanser øger samfundets stabilitet (f.eks. Walker et al., 1999; Pillar et al., 2013). Sammen med den høje grad af både taksonomisk og funktionel overlapning mellem biomerne (figur 1, 5) styrker det faktum, at almindelige, udbredte arter er funktionelt ensartede, desuden forestillingen om, at landplanter på tværs af en bred vifte af miljøforhold deler fælles egenskaber nær kernen af det funktionelle egenskabsspektrum (Wright et al., 2004; Díaz et al., 2016). I modsætning til Umaña et al. (2017) viser vores resultater, at sjældne, geografisk begrænsede arter måske eller måske ikke er funktionelt forskellige fra mere udbredte og almindelige arter. Spørgsmålet er således fortsat åbent, om mere funktionelt særprægede arter er specialister i bestemte miljøer, eller om deres egenskabskombinationer resulterer i demografiske eller fysiologiske kompromiser, der begrænser deres geografiske udbredelse.
Sammenligninger mellem biomer
Vores sammenligninger af egenskabshypervolumenfordelinger på tværs af biomer (figur 2B) er i overensstemmelse med den observation, at mere artsrige miljøer også er mere funktionelt forskelligartede (Swenson et al, 2012; Lamanna et al., 2014; Li et al., 2018; Šímová et al., 2018). Vores resultater er mere tvetydige med hensyn til hypotesen om, at sæsonbestemte og ekstreme klimatiske miljøer konsekvent begrænser arternes funktionelle diversitet (de la Riva et al., 2018). Alle tropiske biomes viser en højere gennemsnitlig funktionel diversitet end alle tempererede biomes, og de polære biomes viser faktisk de mindste hypervolumer. Inden for hver gruppe viser tørre eller mere sæsonmæssigt varierende biomer imidlertid ikke altid mindre hypervolumer (f.eks. tørre skove, xeriske skove), som vi ville have forventet efter tolerancehypotesen af Currie et al. (2004), hvorimod optimale klimatiske forhold understøtter flere kombinationer af fysiologiske parametre. Ud fra disse resultater alene kan vi ikke afgøre, om tempererede og polære biomer er mindre taxonomisk mangfoldige på grund af begrænsninger i det funktionelle nicheområde, eller om deres funktionelle hypervolumer er små, fordi de ikke er taxonomisk mangfoldige. Nulmodelleringstilgange kunne potentielt hjælpe med at adskille taksonomisk og funktionel diversitet (Swenson et al., 2012; Lamanna et al., 2014; Šímová et al., 2018), men en sådan analyse lå uden for rammerne af denne undersøgelse. Mere vigtigt er det, at vores resultater styrker vigtigheden af at forstå, hvordan evolutionær og biogeografisk historie former den funktionelle diversitet af biomers funktionelle diversitet (Woodward et al., 2004; Pennington et al., 2006; Donoghue og Edwards, 2014; Moncrieff et al., 2016). Den omfattende deling af arter (og højere slægtslinjer) på tværs af biomer inden for, men stort set ikke mellem, forskellige biogeografiske områder kunne tjene både til at homogenisere den funktionelle diversitet inden for områderne og til at give ledetråde om de karakteristiske træk, der er udvalgt for eller imod af forskellige miljøer (Douma et al, 2012; Zanne et al., 2014, 2018).
Trods betydelig hypervolumeoverlapning mellem alle biomerne (Supplerende figur 2) synes tropiske, tempererede og kolde biomer alle at besætte adskilte regioner af funktionelt rum (figurer 5B,C). De vigtigste træk, der differentierer biomerne, synes at være træk relateret til den samlede plantestørrelse, herunder både moden højde og frømasse (Supplerende figurer 7-10), snarere end træk vedrørende bladøkonomi, som observeret i mere lokale, plotbaserede analyser (f.eks., Douma et al., 2012). Undtagelsen er blad-P, som viste betydelige forskelle mellem tropiske og tempererede/polære biomer (Supplerende figurer 7-10), et mønster, der er blevet observeret i andre analyser baseret på både artsspecifikke værdier og målinger af hele økosystemer (Kerkhoff et al., 2005; Swenson et al., 2012). Fordi blad P var vores mest sparsomt prøvetagede egenskab, kan forskellene mellem tropiske og tempererede/polare områder være genstand for bias fra imputationsproceduren. Blad P udviser imidlertid et signifikant fylogenetisk signal (Kerkhoff et al., 2006), hvilket tyder på, at imputeringerne bør være ubiased (Swenson et al., 2017). Leaf P er også signifikant højere i urteagtige vækstformer (Kerkhoff et al., 2006), så det latitudinale skift fra overvejende træagtig til urteagtig diversitet (Figur 4) kan også påvirke dette mønster.
Når vi begrænsede hypervolumeanalyser til kun at omfatte de mest udbredte og funktionelt almindelige arter i hvert biome, overlappede de enkelte biomes inden for hvert biogeografisk område mindre i funktionelt egenskabsrum (sammenlign Figur 5B,C), hvilket tyder på, at disse arter kan afspejle fænotyper, der er bedre tilpasset til bestemte miljøer. I denne sammenhæng er det xeriske skovbiom særlig interessant. Delvis på grund af inddragelsen af Chaco-økoregionen er de xeriske skovområder taxonomisk set samlet med de tropiske biomer (figur 5A). Men når man ser på de funktionelle hypervolumer for alle arterne, viser de en langt større lighed med de tempererede biomer (figur 5B). Endelig, når kun de mest udbredte og funktionelt mest almindelige arter analyseres, viser xeriske skovområder igen øget lighed med tropiske biomer, men de opretholder også en høj lighed med tempererede græsarealer og middelhavsbiomerne (figur 5C). Overgange fra varme, mesiske miljøer til koldere, tørre og mere sæsonbestemte miljøer lettes af lignende træk, f.eks. urteagtige habitus (Douma et al., 2012; Zanne et al., 2014, 2018), og ligesom koldere miljøer har samfund i tørre miljøer også en tendens til at være mere fylogenetisk grupperede (Qian og Sandel, 2017). Desuden gør xeriske zoners placering ved grænsen af den intertropiske konvergenszone dem til en geografisk overgang mellem de tropiske og tempererede områder. Det faktum, at xeriske skovområder ligger mellem de tropiske og tempererede områder både funktionelt og biogeografisk, styrker yderligere ideen om, at vi for bedre at forstå biomers funktionelle mangfoldighed må tage hensyn til deres biogeografiske og fylogenetiske historie (Pennington et al., 2006; Donoghue og Edwards, 2014; Moncrieff et al, 2016).
Konklusioner
Alle klassifikationer af terrestriske biomer pålægger et lille antal diskrete kategorier på kontinuerlige gradienter i klima og artsfordelinger, og repræsenterer således en grov forenkling af det komplekse økologiske landskab (Moncrieff et al., 2016, men se Silva de Miranda et al., 2018). Men på trods af deres potentiale for oversimplificering er biomerne nyttige konstruktioner til organisering og forståelse af biodiversiteten og funktionen af Jordens vigtigste terrestriske økosystemer, og trækbaserede tilgange har et stort potentiale til at hjælpe med dynamisk at modellere globale vegetationsfordelinger.
I denne undersøgelse har vi vist, at en stor del af den taksonomiske diversitet i alle biomerne repræsenterer arter, der både er snævert fordelt og funktionelt ens. Inden for biomerne har de mest funktionelt særprægede arter i de enkelte biomer også en tendens til at være geografisk sjældne, mens de udbredte arter ensartet udviser en lav funktionel særprægethed. På trods af omfattende taksonomiske og funktionelle overlapninger mellem biomerne, grupperer de sig i særskilte, biogeografisk og klimatisk adskilte enheder, især når den funktionelle gruppering er baseret på de mest udbredte og funktionelt mest almindelige arter i hvert biome. Fremme af en funktionel forståelse af biomerne vil imidlertid ikke blot kræve en bedre karakterisering af egenskabsvariationen inden for og mellem vegetationstyper, men også oplysninger om den biogeografiske og fylogenetiske historie af artsammensætninger og den relative forekomst af arter inden for biomerne.
Datatilgængelighed
Datasæt af egenskaber og kort over arternes udbredelse, der er analyseret til denne undersøgelse, kan downloades via R-pakken BIEN (Maitner et al., 2018). Se http://bien.nceas.ucsb.edu/bien/ for flere oplysninger om BIEN-databasen.
Author Contributions
SE-L, AK, BJE, DMN og CV designede undersøgelsen; SE-L og AK analyserede dataene; SE-L og AK skrev den første version af manuskriptet; CV, BB, NK, BSM, BM, RKP, BS, SS, J-CS og SKW bidrog til indsamling og oprettelse af egenskabs- og rækkeviddekortdatabasen. Alle forfattere bidrog til udvikling og skrivning af manuskriptet.
Funding
SE-L, DMN og AK blev støttet af en samarbejdsbevilling til forskningssamarbejde fra US National Science Foundation (DEB-1556651). BJE blev støttet af National Science Foundation-pris DEB-1457812 og Macrosystems-1065861. CV blev støttet af Det Europæiske Forskningsråd (Grant ERC-StG-2014-639706-CONSTRAINTS) og af den franske fond for forskning i biodiversitet (FRB; www.fondationbiodiversite.fr) i forbindelse med CESAB-projektet Årsager og konsekvenser af funktionel sjældenhed fra lokale til globale skalaer. J-CS betragter dette arbejde som et bidrag til sit VILLUM Investigator-projekt Biodiversity Dynamics in a Changing World, der finansieres af VILLUM FONDEN (bevilling 16549). SKW blev støttet af Strategic Science Investment Fund fra New Zealand Ministry of Business, Innovation and Employment’s Science and Innovation Group.
Interessekonflikterklæring
Forfatterne erklærer, at forskningen blev udført uden kommercielle eller finansielle relationer, der kunne opfattes som en potentiel interessekonflikt.
Akkommentarer
Vi takker Dan Griffith og to korrekturlæsere for deres konstruktive kritik, som i høj grad forbedrede dette manuskript. Vi takker også alle bidragydere af BIEN-data (se http://bien.nceas.ucsb.edu/bien/data-contributors/ for en komplet liste).
Supplementært materiale
Det supplerende materiale til denne artikel kan findes online på: https://www.frontiersin.org/articles/10.3389/fevo.2018.00219/full#supplementary-material
Banda, R.K., Delgado-Salinas, A., Dexter, K. G., Linares-Palomino, R., Oliveira-Filho, A., Prado, D., et al. (2016). Plantediversitetsmønstre i neotropiske tørre skove og deres konsekvenser for bevarelsen. Science 353, 1383-1387. doi: 10.1126/science.aaf5080
PubMed Abstract | CrossRef Full Text | Google Scholar
Blonder, B., Lamanna, C., Lamanna, C., Violle, C., and Enquist, B. J. (2014). Den n-dimensionelle hypervolume. Glob. Ecol. Biogeogr. 23, 595-609. doi: 10.1111/geb.12146
CrossRef Full Text | Google Scholar
Blonder, B., Morrow, C. B., Maitner, B., Harris, D. J., Lamanna, C., Violle, C., et al. (2018). Nye tilgange til afgrænsning af n-dimensionelle hypervolumer. Methods Ecol. Evol. 9, 305-319. doi: 10.1111/2041-210X.12865
CrossRef Full Text | Google Scholar
Bonan, G. B., Oleson, K. W., Fisher, R. A., Lasslop, G., og Reichstein, M. (2012). Afstemning af bladfysiologiske egenskaber og data om kronedækningsstrømme: anvendelse af TRY- og FLUXNET-databaserne i Fællesskabets landmodel version 4. J. Geophys. Res. Biogeosci. 117:G02026. doi: 10.1029/2011JG001913
CrossRef Full Text | Google Scholar
Borregaard, M. K., og Rahbek, C. (2010). Kausalitet i forholdet mellem geografisk udbredelse og artshyppighed. Q. Rev. Biol. 85, 3-25. doi: 10.1086/650265
PubMed Abstract | CrossRef Full Text | Google Scholar
Bruggeman, J., Heringa, J., og Brandt, B. W. (2009). PhyloPars: estimering af manglende parameterværdier ved hjælp af fylogeni. Nucleic Acids Res. 37, W179-W184. doi: 10.1093/nar/gkp370
PubMed Abstract | CrossRef Full Text | Google Scholar
Cadotte, M. W., Carscadden, K., og Mirotchnick, N. (2011). Ud over arter: funktionel mangfoldighed og opretholdelse af økologiske processer og tjenester. J. Appl. Ecol. 48, 1079-1087. doi: 10.1111/j.1365-2664.2011.02048.x
CrossRef Full Text | Google Scholar
Cornwell, W. K., Cornelissen, J. H., Amatangelo, K., Dorrepaal, E., Eviner, V. T., Godoy, O., et al. (2008). Plantearternes egenskaber er den fremherskende kontrol for nedbrydningshastigheden af affald inden for biomer verden over. Ecol. Lett. 11, 1065-1071. doi: 10.1111/j.1461-0248.2008.01219.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Craine, J. M. (2009). Ressourcestrategier for vilde planter. New Jersey, NJ: Princeton University Press. doi: 10.1515/9781400830640
CrossRef Full Text | Google Scholar
Currie, D. J., Mittelbach, G. G. G., Cornell, H. V., Field, R., Guégan, J. F., Hawkins, B. A., et al. (2004). Predictions and tests of climate-based hypotheses of broad-scale variation in taxonomic richness (Forudsigelser og test af klimabaserede hypoteser om variation i bred skala i taksonomisk rigdom). Ecol. Lett. 12, 1121-1134. doi: 10.1111/j.1461-0248.2004.00671.x
CrossRef Full Text | Google Scholar
de la Riva, E. G., Violle, C., Pérez-Ramos, I. M., Marañón, T., Navarro-Fernández, C. M., Olmo, M., et al. (2018). En multidimensionel funktionel egenskabstilgang afslører aftryk af miljøstress i Middelhavets træagtige Fællesskaber. Ecosystems 21, 248-262. doi: 10.1007/s10021-017-0147-7
CrossRef Full Text | Google Scholar
Díaz, S., Kattge, J., Cornelissen, J. H., Wright, I. J., Lavorel, S., Dray, S., et al. (2016). Det globale spektrum af planters form og funktion. Nature 529, 167-171. doi: 10.1038/nature16489
PubMed Abstract | CrossRef Full Text | Google Scholar
Díaz, S., Lavorel, S., de Bello, F., Quétier, F., Grigulis, K., og Robson, T. M. (2007). Indarbejdelse af virkninger af plantefunktionel diversitet i vurderinger af økosystemtjenester. Proc. Natl. Acad. Sci. U.S.A 104, 20684-20689. doi: 10.1073/pnas.0704716104
PubMed Abstract | CrossRef Full Text | Google Scholar
Donoghue, M. J., og Edwards, E. J. (2014). Biome-skift og niche-evolution hos planter. Annu. Rev. Ecol. Evol. syst. 45, 547-572. doi: 10.1146/annurev-ecolsys-120213-091905
CrossRef Full Text | Google Scholar
Douma, J. C., Aerts, R., Witte, J. P. M. M., Bekker, R. M., Kunzmann, D., Metselaar, K., et al. (2012). En kombination af funktionelt forskellige planteegenskaber giver mulighed for kvantitativt at forudsige en bred vifte af artsammensætninger i det nordvestlige Europa. Ecography 35, 364-373. doi: 10.1111/j.1600-0587.2011.07068.x
CrossRef Full Text | Google Scholar
Engemann, K., Sandel, B., Enquist, B. J. J., Jørgensen, P. M., Kraft, N., Marcuse-Kubitza, A., et al. (2016). Mønstre og drivkræfter for dominans af plantefunktionelle grupper på tværs af den vestlige halvkugle: en makroøkologisk revurdering baseret på et massivt botanisk datasæt. Bot. J. Linn. Soc. 180, 141-160. doi: 10.1111/boj.12362
CrossRef Full Text | Google Scholar
Enquist, B. J., Condit, R., Peet, R., Thiers, B., og Schildhauer, M. (2016). Det botaniske informations- og økologinetværk (BIEN): cyberinfrastruktur for et integreret botanisk informationsnetværk til undersøgelse af de økologiske virkninger af globale klimaændringer på plantebiodiversiteten. PeerJ Prepr. 4:e2615v2. doi: 10.7287/peerj.preprints.2615v2
CrossRef Full Text | Google Scholar
Garnier, E., Cortez, J., Billès, G., Navas, M. L., Roumet, C., Debussche, M., et al. (2004). Plantefunktionelle markører fanger økosystemets egenskaber under sekundær succession. Ecology 85, 2630-2637. doi: 10.1890/03-0799
CrossRef Full Text | Google Scholar
Goldsmith, G. R., Morueta-Holme, N., Sandel, B., Fitz, E. D., Fitz, S. D., Boyle, B., et al. (2016). Plant-O-Matic: en dynamisk og mobil guide til alle planter i Amerika. Metoder Ecol. Evol. 7, 960-965. doi: 10.1111/2041-210X.12548
CrossRef Full Text | Google Scholar
Goolsby, E. W., Bruggeman, J., and Ané, C. (2017). Rphylopars: hurtige multivariate fylogenetiske komparative metoder til manglende data og variation inden for arten. Methods Ecol. Evol. 8, 22-27. doi: 10.1111/2041-210X.12612
CrossRef Full Text | Google Scholar
Grenié, M., Denelle, P., Tucker, C. M., Munoz, F., og Violle, C. (2017). funrar: an R package to characterize functional rarity. Divers. Distrib. 23, 1365-1371. doi: 10.1111/ddi.12629
CrossRef Full Text | Google Scholar
Hortal, J., de Bello, F., Diniz-Filho, J. A. F., Lewinsohn, T. M., Lobo, J. M., og Ladle, R. J. (2015). Syv mangler, der plager viden om biodiversitet i stor skala. Annu. Rev. Ecol. Evol. Syst. 46, 523-549. doi: 10.1146/annurev-ecolsys-112414-054400
CrossRef Full Text | Google Scholar
Kerkhoff, A. J., Enquist, B. J., Enquist, B. J., Elser, J. J., og Fagan, W. F. (2005). Planteallometri, stoichiometri og temperaturafhængighed af primær produktivitet. Glob. Ecol. Biogeogr. 14, 585-598. doi: 10.1111/j.1466-822X.2005.00187.x
CrossRef Full Text | Google Scholar
Kerkhoff, A. J., Fagan, W. F., Elser, J. J., og Enquist, B. J. (2006). Fylogenetisk og vækstformsvariation i skalering af kvælstof og fosfor i frøplanter. Am. Nat. 168, E103-E122. doi: 10.1086/507879
PubMed Abstract | CrossRef Full Text | Google Scholar
Kraft, N. J., Valencia, R., og Ackerly, D. D. (2008). Funktionelle træk og nichebaseret træfællesskabssammensætning i en amazonisk skov. Science 322, 580-582. doi: 10.1126/science.1160662
PubMed Abstract | CrossRef Full Text | Google Scholar
Lamanna, C., Blonder, B., Violle, C., Kraft, N. J., Sandel, B., et al. (2014). Funktionelt trækrum og den latitudinale diversitetsgradient. Proc. Natl. Acad. Sci. U.S.A. 111, 13745-13750. doi: 10.1073/pnas.1317722111
PubMed Abstract | CrossRef Full Text | Google Scholar
Lavorel, S., og Garnier, E. (2002). Forudsigelse af ændringer i samfundssammensætningen og økosystemets funktion ud fra planteegenskaber: en fornyet gennemgang af den hellige gral. Funct. Ecol. 16, 545-556. doi: 10.1046/j.1365-2435.2002.00664.x
CrossRef Full Text | Google Scholar
Lavorel, S., Grigulis, K., Lamarque, P., Colace, M.-P., Garden, D., Girel, J., et al. (2011). Brug af funktionelle planteegenskaber til at forstå landskabsfordelingen af flere økosystemtjenester. J. Ecol. 99, 135-147. doi: 10.1111/j.1365-2745.2010.01753.x
CrossRef Full Text | Google Scholar
Li, Y., Shipley, B., Price, J. N., Dantas, V. de L., Tamme, R., Westoby, M., et al. (2018). Habitatfiltrering bestemmer den funktionelle nichebesættelse af plantesamfund i hele verden. J. Ecol. 106, 1001-1009. doi: 10.1111/1365-2745.12802
CrossRef Full Text | Google Scholar
Magallón, S., Gómez-Acevedo, S., Sánchez-Reyes, L. L. L., og Hernández-Hernández, T. (2015). Et metakalibreret tidstræ dokumenterer den tidlige stigning i blomstrende planters fylogenetiske diversitet. New Phytol. 207, 437-453. doi: 10.1111/nph.13264
PubMed Abstract | CrossRef Full Text | Google Scholar
Maitner, B. S., Boyle, B., Casler, N., Condit, R., Donoghue, J., Durán, S. M., et al. (2018). BIEN R-pakken: et værktøj til at få adgang til databasen Botanical Information and Ecology Network (BIEN). Methods Ecol. Evol. 9, 373-379. doi: 10.1111/2041-210X.12861
CrossRef Full Text | Google Scholar
Moncrieff, G. R., Bond, W. J., og Higgins, S. I. (2016). Revising the biome concept for understanding and predicting global change impacts. J. Biogeogr. 43, 863-873. doi: 10.1111/jbi.12701
CrossRef Full Text | Google Scholar
Novosolov, M., Rodda, G. H., North, A. C., Butchart, S. H. M., Tallowin, O. J. S., Gainsbury, A. M., et al. (2017). Populationstæthed-range size relationship revisited. Glob. Ecol. Biogeogr. 26, 1088-1097. doi: 10.1111/geb.12617
CrossRef Full Text | Google Scholar
Olson, D. M., Dinerstein, E., Wikramanayake, E. D., Burgess, N. D., Powell, G. V. N., Underwood, E. C., et al. (2001). Terrestrial Ecoregions of the world: a new map of life on earth: a new global map of terrestrial ecoregions provides an innovative tool for conserving biodiversity. Bioscience 51, 933-938.doi: 10.1641/0006-3568(2001)0512.0.CO;2
CrossRef Full Text | Google Scholar
Pennington, R. T., Prado, D. E., and Pendry, C. A. (2000). Neotropiske sæsonbestemte tørre skove og kvartære vegetationsændringer. J. Biogeogr. 27, 261-273. doi: 10.1046/j.1365-2699.2000.00397.x
CrossRef Full Text | Google Scholar
Pennington, R. T., Richardson, J. E., and Lavin, M. (2006). Indsigt i den historiske opbygning af artsrige biomes fra daterede plantefylogenier, neutral økologisk teori og fylogenetisk samfundsstruktur. New Phytol. 172, 605-616. doi: 10.1111/j.1469-8137.2006.01902.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Penone, C., Davidson, A. D., Shoemaker, K. T., Di Marco, M., Rondinini, C., Brooks, T. M., et al. (2014). Imputation af manglende data i datasæt med livshistoriske egenskaber: Hvilken tilgang er den bedste? Methods Ecol. Evol. 5, 961-970. doi: 10.1111/2041-210X.12232
CrossRef Full Text | Google Scholar
Pillar, V. D., Blanco, C. C. C., Müller, S. C., Sosinski, E. E., Joner, F., og Duarte, L. D. S. (2013). Funktionel redundans og stabilitet i plantesamfund. J. Veg. Sci. 24, 963-974. doi: 10.1111/jvs.12047
CrossRef Full Text | Google Scholar
Prado, D. E., og Gibbs, P. E. (1993). Mønstre af artsfordelinger i tørre sæsonbestemte skove i Sydamerika. Ann. Mo. Bot. Gard. 80, 902-927. doi: 10.2307/2399937
CrossRef Full Text | Google Scholar
Qian, H., and Sandel, B. (2017). Fylogenetisk struktur af regionale angiosperm-samlinger på tværs af bredde- og klimatiske gradienter i Nordamerika. Glob. Ecol. Biogeogr. 26, 1258-1269. doi: 10.1111/geb.12634
CrossRef Full Text | Google Scholar
Reich, P. B., Wright, I. J., Cavender-Bares, J., Craine, J. M., Oleksyn, J., Westoby, M., et al. (2003). Udviklingen af funktionel variation i planter: træk, spektrer og strategier. Int. J. Plant Sci. 164, S143-S164. doi: 10.1086/374368
CrossRef Full Text | Google Scholar
Silva de Miranda, P. L., Oliveira-Filho, A., Pennington, R. T., Neves, D. M., Baker, T. R., og Dexter, K. G. (2018). Brug af træartsopgørelser til at kortlægge biomes og vurdere deres klimatiske overlapninger i lavlandets tropiske Sydamerika. Glob. Ecol. Biogeogr. 27, 899-912. doi: 10.1111/geb.12749
CrossRef Full Text | Google Scholar
Šímová, I., Violle, C., Kraft, N. J. B. B., Storch, D., Svenning, J.-C., Boyle, B., et al. (2015). Forskydninger i egenskabsmiddelværdier og varianser i nordamerikanske træforsamlinger: Artsrigdomsmønstre er løst relateret til det funktionelle rum. Ecography 38, 649-658. doi: 10.1111/ecog.00867
CrossRef Full Text | Google Scholar
Šímová, I., Violle, C., Svenning, J.-C., Kattge, J., Engemann, K., Sandel, B., et al. (2018). Rumlige mønstre og klimaforhold for vigtige planteegenskaber i den nye verden adskiller sig mellem træagtige og urteagtige arter. J. Biogeogr. 45, 895-916. doi: 10.1111/jbi.13171
CrossRef Full Text | Google Scholar
Smith, S. A., og Beaulieu, J. M. (2009). Livshistorie påvirker hastighederne for klimatisk nicheudvikling hos blomstrende planter. Proc. Biol. Sci. 276, 4345-4352. doi: 10.1098/rspb.2009.1176
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, S. A., og Brown, J. W. (2018). Konstruktion af en bredt inkluderende frøplantefylogeni. Am. J. Bot. 105, 302-314. doi: 10.1002/ajb2.1019
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, S. A., og Donoghue, M. J. (2008). Hastighederne for molekylær evolution er knyttet til livshistorien hos blomstrende planter. Science 322, 86-89. doi: 10.1126/science.1163197
PubMed Abstract | CrossRef Full Text | Google Scholar
Swenson, N. G. (2014). Phylogenetic imputation of plant functional trait databases. ecography 37, 105-110. doi: 10.1111/j.1600-0587.2013.00528.x
CrossRef Full Text
Swenson, N. G., Enquist, B. J., Pither, J., Kerkhoff, A. J., Boyle, B., Weiser, M. D., et al. (2012). Biogeografi og filtrering af funktionel diversitet af træagtige planter i Nord- og Sydamerika. Glob. Ecol. Biogeogr. 21, 798-808. doi: 10.1111/j.1466-8238.2011.00727.x
CrossRef Full Text | Google Scholar
Swenson, N. G., Weiser, M. D., Mao, L., Araújo, M. B., Diniz-Filho, J. A. F., Kollmann, J., et al. (2017). Fylogeni og forudsigelse af træernes funktionelle diversitet på tværs af nye kontinentale indstillinger. Glob. Ecol. Biogeogr. 26, 553-562. doi: 10.1111/geb.12559
CrossRef Full Text | Google Scholar
Umaña, M. N., Mi, X., Cao, M., Enquist, B. J., Hao, Z., Howe, R., et al. (2017). Den rolle, som funktionel entydighed og rumlig aggregering spiller i forklaringen af sjældenhed i træer. Glob. Ecol. Biogeogr. 26, 777-786. doi: 10.1111/geb.12583
CrossRef Full Text | Google Scholar
van Bodegom, P. M., Douma, J. C., og Verheijen, L. M. (2014). En fuldt egenskabsbaseret tilgang til modellering af global vegetationsfordeling. Proc. Natl. Acad. Sci. U.S.A. 111, 13733-13738. doi: 10.1073/pnas.1304551110
PubMed Abstract | CrossRef Full Text | Google Scholar
Violle, C., Reich, P. B., Pacala, S. W., Enquist, B. J., og Kattge, J. (2014). Fremkomsten og løftet af funktionel biogeografi. Proc. Natl. Acad. Sci. U.S.A. 111, 13690-13696. doi: 10.1073/pnas.1415442111
PubMed Abstract | CrossRef Full Text | Google Scholar
Violle, C., Thuiller, W., Mouquet, N., Munoz, F., Kraft, N. J. B., Cadotte, M. W., et al. (2017). Funktionel sjældenhed: økologi af outliers. Trends Ecol. Evol. 32, 356-367. doi: 10.1016/j.tree.2017.02.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Walker, B., Kinzig, A., og Langridge, J. (1999). Planteattediversitet, modstandsdygtighed og økosystemfunktion: karakteren og betydningen af dominerende og mindre betydningsfulde arter. Ecosystems 2, 95-113. doi: 10.1007/s100219900062
CrossRef Full Text | Google Scholar
Westoby, M. (1998). Et planteøkologisk strategisystem for bladhøjde-sæd (LHS). Plant Soil 199, 213-227. doi: 10.1023/A:1004327224729
CrossRef Full Text | Google Scholar
Woodward, F. I., Lomas, M. R., og Kelly, C. K. (2004). Globalt klima og fordelingen af plantebiomer. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 359, 1465-1476. doi: 10.1098/rstb.2004.1525
PubMed Abstract | CrossRef Full Text | Google Scholar
Wright, I. J., Reich, P. B., Westoby, M., Ackerly, D. D., Baruch, Z., Bongers, F., et al. (2004). Det verdensomspændende bladøkonomiske spektrum. Nature 428, 821-827. doi: 10.1038/nature02403
PubMed Abstract | CrossRef Full Text | Google Scholar
Xia, J., Niu, S., Ciais, P., Janssens, I. A., Chen, J., Ammann, C., et al. (2015). Fælles kontrol af den terrestriske bruttoprimærproduktivitet ved hjælp af planters fænologi og fysiologi. Proc. Natl. Acad. Sci. U.S.A. 112, 2788-2793. doi: 10.1073/pnas.1413090112
PubMed Abstract | CrossRef Full Text | Google Scholar
Zanne, A. E., Pearse, W. D., Cornwell, W. K., McGlinn, D. J., Wright, I. J., og Uyeda, J. C. (2018). Funktionel biogeografi af angiospermer: liv ved ekstremerne. New Phytol. 218, 1697-1709. doi: 10.1111/nph.15114
PubMed Abstract | CrossRef Full Text | Google Scholar
Zanne, A. E., Tank, D. C., Cornwell, W. K., Eastman, J. M., Smith, S. A., FitzJohn, R. G., et al. (2014). Tre nøgler til udstråling af angiospermer til frysemiljøer. Nature 506, 89-92. doi: 10.1038/nature12872
PubMed Abstract | CrossRef Full Text | Google Scholar