Vurdering | Biopsykologi | Komparativ | Kognitiv | Udvikling | Sprog | Individuelle forskelle | Personlighed | Filosofi | Social |
Metoder | Statistik | Klinisk | Pædagogisk | Industriel | Professionelle emner | Verdenspsykologi |
Biologisk:Adfærdsgenetik – Evolutionær psykologi – Neuroanatomi – Neurokemi – Neuroendokrinologi -Neurovidenskab – Psykoneuroimmunologi – Fysiologisk psykologi – Psykofarmakologi(Indeks, Oversigt)
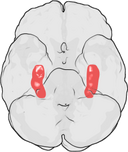
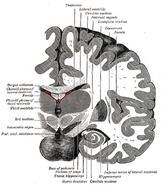
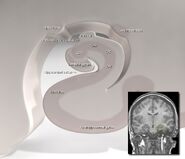
Hippocampus’ placering i den menneskelige hjerne.
For tidsskriftet af samme navn, se Hippocampus (tidsskrift)
Hippocampus er en del af forhjernen, der er placeret i den mediale tindingelap. Den hører til det limbiske system og spiller vigtige roller i langtidshukommelsen og den rumlige navigation. Mennesker og andre pattedyr har to hippocampi, en i hver side af hjernen. Hos gnavere, hvor den er blevet undersøgt mest indgående, er hippocampus formet som en banan. Hos mennesker har den en buet og snoet form, som mindede de tidlige anatomikere om en søhest. Navnet stammer faktisk fra det græske ord for søhest (græsk: hippos = hest, campos = hav).
I Alzheimers sygdom er hippocampus en af de første regioner i hjernen, der lider skade; hukommelsesproblemer og desorientering er blandt de første symptomer. Skader på hippocampus kan også skyldes iltmangel (anoxi), hjernebetændelse eller epilepsi i mesiale tindingelapper. Personer med omfattende hippocampusskader kan opleve amnesi, dvs. manglende evne til at danne eller fastholde nye erindringer.
Hippocampus’ funktioner
Måske var den tidligste idé, at hippocampus er involveret i lugtesansen: dette synes primært at være blevet antydet på grund af dens placering i hjernen, ved siden af den olfaktoriske cortex. Der er fortsat en vis interesse for hippocampus’ olfaktoriske responser, men næsten ingen mener nu, at hippocampus’ primære funktion er olfaktorisk.
I årenes løb har tre hovedidéer om hippocampus’ funktion domineret litteraturen: hæmning, hukommelse og rum. Teorien om adfærdsmæssig hæmning (karikeret af O’Keefe og Nadel som “træd på bremsen!”) var meget populær op til 1960’erne. Den fik meget af sin styrke fra to observationer: for det første har dyr med hippocampusskader tendens til at være hyperaktive, og for det andet har dyr med hippocampusskader ofte svært ved at lære at hæmme reaktioner, som de tidligere har lært. Jeffrey Gray udviklede denne tankegang til en fuldgyldig teori om hippocampus’ rolle i forbindelse med angst. Hæmningsteorien er dog ikke særlig populær på nuværende tidspunkt.
Den anden vigtige tankegang relaterer hippocampus til hukommelse. Selv om den havde forløbere, fik denne idé sin hovedkraft fra en meget velkendt rapport af Scoville og Milner om resultaterne af en kirurgisk ødelæggelse af hippocampus (i et forsøg på at lindre epileptiske anfald) hos en patient kendt som H.M. Det uventede resultat var alvorlig hukommelsestab: H.M. var ude af stand til bevidst at huske begivenheder, der fandt sted efter hans operation eller i flere år før den. Dette tilfælde vakte så stor interesse, at H.M. nu siges at være det mest intensivt undersøgte medicinske tilfælde i historien. I de efterfølgende år er andre patienter med lignende niveauer af hippocampusskader og hukommelsestab (forårsaget af ulykker eller sygdom) også blevet undersøgt, og bogstavelig talt tusindvis af eksperimenter har undersøgt fysiologien bag neuronal plasticitet i hippocampus. Der er nu næsten universel enighed om, at hippocampus spiller en eller anden form for vigtig rolle for hukommelsen; den præcise karakter af denne rolle er dog stadig meget omdiskuteret.
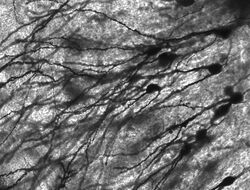
Golgi-farvede neuroner i gnaverens hippocampus.
Rolle i hukommelsen
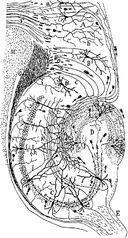
Tegning af det neurale kredsløb i gnaverens hippocampus. S. Ramón y Cajal, 1911.
Psykologer og neurovidenskabsfolk er uenige om hippocampus’ præcise rolle, men er generelt enige om, at den spiller en afgørende rolle i dannelsen af nye erindringer om personligt oplevede begivenheder (episodisk eller selvbiografisk hukommelse). Nogle forskere foretrækker at betragte hippocampus som en del af et større hukommelsessystem i den mediale tindingelap, der er ansvarlig for generel deklarativ hukommelse (erindringer, der kan verbaliseres eksplicit – disse ville for eksempel omfatte hukommelse for fakta ud over episodisk hukommelse).
Hovedartikel: Hippocampus’ rolle i hukommelse
En del beviser implicerer hippocampus i lagring og behandling af rumlig information. Undersøgelser på rotter har vist, at neuroner i hippocampus har rumlige fyringsfelter. Disse celler kaldes stedceller. Nogle celler fyrer, når dyret befinder sig på et bestemt sted, uanset bevægelsesretning, mens de fleste er i det mindste delvist følsomme over for hovedets retning og bevægelsesretning.
Hovedartikel: Hippocampus’ rolle i rumlig hukommelse og navigation
Hippocampus og depression
- Tab af hippocampal volumen, der måske afspejler virkningerne af overdrevne glukokortikoider på
neurogenese, er blevet identificeret i grupper af deprimerede mennesker i sammenligning med kontrolpersoner.
Hovedartikel: Depression og hippocampus
Historie
Anatomen Giulio Cesare Aranzi (ca. 1564) brugte først betegnelsen hippocampus til at beskrive det cerebrale organ på grund af dets visuelle lighed med en søhest. Dette organ blev oprindeligt forbundet med lugtesansen snarere end med dets kendte funktion i forbindelse med erhvervelse af hukommelse. Russeren Vladimir Bekhterev bemærkede hippocampus’ rolle i hukommelsen omkring 1900 på baggrund af observationer af en patient med alvorlige hukommelsesforstyrrelser. I mange år var den konventionelle opfattelse af hippocampus imidlertid, at den ligesom resten af det limbiske system var ansvarlig for følelser.
Hippocampus’ betydning for hukommelsen blev gjort opmærksom på forskerne af patienten HM. HM led af en række anterograde og tidsmæssigt graduerede retrograde hukommelsesforstyrrelser (sådanne forstyrrelser er emnet for filmen Memento) efter bilateral fjernelse af forskellige strukturer i den medial-temporale lobe (herunder bilateral ablation af hans hippocampi) for at lindre hyppige epileptiske anfald. Af særlig betydning er det, at HM stadig var i stand til at lære proceduremæssige opgaver (som er forbundet med striatum) og havde en IQ over gennemsnittet. HM udviste en slående enkelt-dissociation mellem intelligens og deklarativ hukommelse. Den relative størrelse af hippocampusformationen i forhold til hjernens samlede volumen er ofte bevaret hos de fleste pattedyrsarter. Ikke desto mindre har det vist sig, at disse områder er relativt hypotrofiske hos hvaler.
Anatomi
Hovedartikel: Hippocampus anatomi
Humane hippocampus.
Nissl-farvet koronalt snit af hjernen fra en makakakabe, der viser hippocampus (indcirklet). Kilde: brainmaps.org
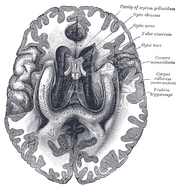
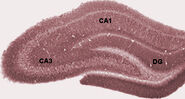
Anatomisk set er hippocampus en uddybning af kanten afcortex. Den kan skelnes som en zone, hvor cortexen indsnævres til et enkelt lag af meget tætpakkede neuroner, som krøller sig sammen i en stram S-form. De strukturer, der ligger langs cortexens kant, udgør det såkaldte limbiske system (latin limbus = kant): disse omfatter hippocampus, cingulatcortex, olfaktoriskcortex og amygdala. Paul MacLean foreslog engang, som en del af sin teori om den tredelte hjerne, at de limbiske strukturer omfatter det neurale grundlag for følelser. De fleste neurovidenskabsfolk mener dog ikke længere, at begrebet om et samlet “limbisk system” er gyldigt.
Hippocampus som helhed ender med at ligne et buet rør, som er blevet analogiseret med en søhest, et vædderhorn (Cornu Ammonis) eller en banan.
Dette generelle layout gælder for alle pattedyrarter, fra pindsvin til menneske, selv om detaljerne varierer. Hos rotten ligner de to hippocampi på forbløffende vis et par bananer, der er forbundet ved stilken. I menneske- og abehjerner er den nederste del af hippocampus i bunden af hjernen, tæt på tindingelappens basis, meget bredere end den øverste del. En af konsekvenserne af denne komplekse geometri er, at tværsnit gennem hippocampus kan vise en forvirrende variation af former, afhængigt af snitets vinkel og placering.
Hippocampus’ stærkeste forbindelser er med den entorhinale cortex (EC), som ligger ved siden af den i tindingelappen. De overfladiske lag af EC giver de mest talrige input til hippocampus, og de dybe lag af EC modtager de mest talrige outputs. EC er til gengæld stærkt og gensidigt forbundet med mange andre dele af hjernebarken. Hippocampus modtager også en meget vigtig projektion fra det mediale septumområde. Ødelæggelse af det septale område ophæver den hippocampale theta-rytme og forringer visse typer hukommelse alvorligt. (Såkaldte “date rape”-stoffer menes at udøve deres hukommelseshæmmende virkninger i det mindste delvist ved at modvirke den kolinerge projektion fra det mediale septum til hippocampus.)
Fysiologi
Eksempler på rottehippocampal EEG og CA1-neuroaktivitet i theta- (vågen/opvækket) og LIA- (slow-wave sleep) tilstand. Hvert plot viser 20 sekunders data med et hippocampal EEG-spor øverst, spike-raster fra 40 samtidig registrerede CA1-pyramidale celler i midten (hver rasterlinje repræsenterer en anden celle) og et plot af løbehastigheden nederst. Det øverste plot repræsenterer en tidsperiode, hvor rotten aktivt søgte efter spredte fødekugler. I det nederste plot sov rotten.
Hippocampus viser to store “tilstande” af aktivitet, som hver især er forbundet med et særskilt mønster af EEG-bølger og neurale populationsaktiviteter.Disse tilstande er opkaldt efter de EEG-mønstre, der er forbundet med dem: theta og stor uregelmæssig aktivitet (LIA). Her er nogle af deres vigtigste karakteristika hos rotten, som er det dyr, der er blevet studeret mest omfattende:
Theta-tilstanden optræder under aktiv, vågen adfærd (især ved bevægelse) og også under REM-søvn (drømme). I theta-tilstanden er EEG’et domineret af store regelmæssige bølger med et frekvensområde på 6-9 Hz, og de vigtigste grupper af hippocampale neuroner (pyramideceller og granuleceller) udviser sparsom populationsaktivitet, hvilket betyder, at i ethvert kort tidsinterval er langt størstedelen af cellerne tavse, mens den lille resterende fraktion affyres med forholdsvis høj hastighed, op til 50 spikes i et sekund for de mest aktive af dem. En aktiv celle forbliver typisk aktiv i mellem et halvt sekund og et par sekunder. Efterhånden som rotten opfører sig, bliver de aktive celler tavse, og nye celler bliver aktive, men den samlede procentdel af aktive celler forbliver mere eller mindre konstant. I mange situationer bestemmes cellaktiviteten i høj grad af dyrets geografiske placering, men andre adfærdsvariabler har også en klar indflydelse på den.
LIA-tilstanden optræder under langsom bølgesøvn (ikke-drømmende) og også under tilstande af immobilitet i vågen tilstand, f.eks. når man hviler eller spiser. I LIA-tilstanden domineres EEG’et af skarpe bølger, som er tilfældigt timede store afbøjninger af EEG-signalet, der varer i 200-300 sek. Disse skarpe bølger bestemmer også de neurale aktivitetsmønstre i befolkningen. Mellem dem er pyramidale celler og granuleceller meget stille (men ikke tavse). Under en skarp bølge kan så mange som 5-10% af populationen udsende aktionspotentialer i løbet af en periode på 50 msek; mange af disse celler udsender ikke én, men et burst af spikes.
Disse to hippocapampale aktivitetsformer kan ses hos primater såvel som hos rotter, med den vigtige undtagelse, at det har været vanskeligt at se en robust theta-rytmicitet i hippocampus hos primater. Der er dog kvalitativt set ens skarpe bølger og lignende statsafhængige ændringer i den neurale befolkningsaktivitet.
Thetarytmen
På grund af sine tætpakkede neurale lag genererer hippocampus nogle af de største EEG-signaler af alle hjernestrukturer. I nogle situationer er EEG’et domineret af regelmæssige bølger, der ofte fortsætter i mange sekunder. Dette EEG-mønster er kendt som thetarytme. Det var et af de tidligste EEG-fænomener, der blev opdaget: den første beskrivelse kom fra Jung og Kornmuller i 1938. Det var imidlertid først i 1954, da Green og Arduini offentliggjorde en lang og grundig undersøgelse af thetarytmen hos kaniner, katte og aber, at interessen for alvor tog fart, og måske især fordi de satte thetarytmen i forbindelse med ophidselse, som var tidens aktuelle emne, udløste deres artikel en strøm af opfølgende undersøgelser, hvilket resulterede i bogstaveligt talt hundredvis af undersøgelser af thetarytmens fysiologi og farmakologi i løbet af 1950’erne og 1960’erne. På trods af dette ret skræmmende arbejde forblev mange spørgsmål ubesvarede, især spørgsmålet om funktion. Selv på nuværende tidspunkt er dette mest kritiske spørgsmål endnu ikke blevet besvaret på overbevisende vis.
Theta-rytmik er meget tydelig hos kaniner og gnavere, og den er også tydelig hos katte og hunde. Hvorvidt theta kan ses hos primater er et irriterende spørgsmål. Green og Arduini rapporterede kun om meget korte udbrud af temmelig uregelmæssig rytmicitet hos aber, og de fleste senere undersøgelser har ikke set meget mere. Variationer i metodologien har imidlertid gjort det vanskeligt at drage stærke konklusioner.
I rotter (de dyr, der er blevet undersøgt langt mest indgående) ses theta hovedsageligt under to forhold: for det første, når et dyr går eller på anden måde interagerer aktivt med omgivelserne, og for det andet under REM-søvn. Frekvensen stiger som en funktion af løbehastigheden, idet den starter ved ca. 6,5 Hz i den lave ende og stiger til ca. 9 Hz i den høje ende, selv om der undertiden ses højere frekvenser ved dramatiske bevægelser, f.eks. ved spring over store kløfter. Hos andre, større dyrearter er thetafrekvenserne generelt en smule lavere. Adfærdsafhængigheden synes også at variere fra art til art: hos katte og kaniner observeres theta ofte under tilstande af ubevægelig årvågenhed. Dette er også blevet rapporteret for rotter, men kun når de er alvorligt skræmte.
Theta er ikke kun begrænset til hippocampus. Hos rotter kan det iagttages i mange dele af hjernen, herunder næsten alle, der interagerer stærkt med hippocampus. Man mener, at pacemakeren for rytmen ligger i det mediale septumområde: dette område projicerer til alle de regioner, der viser theta-rytme, og ødelæggelse af dette område eliminerer theta i hele hjernen. (Der kan være én undtagelse, nemlig et lille område i hypothalamus kaldet supramamillarynucleus, som synes at være i stand til at opretholde theta uafhængigt af septum i nogle situationer.)
Thetas funktion, hvis man antager, at den har en, er endnu ikke blevet forklaret på overbevisende vis, selv om der er blevet foreslået mange teorier.Den mest populære tendens har været at relatere den til indlæring og hukommelse.Det er veletableret, at læsioner af det mediale septum–det centrale knudepunkt i thetasystemet–forårsager alvorlige forstyrrelser af hukommelsen.Det mediale septum er imidlertid mere end blot kontrollanten af theta,det er også hovedkilden til kolinerge projektioner tilhippocampus. Det er ikke blevet fastslået, at septallæsioner udøver deres virkninger specifikt ved at eliminere theta.
Skarpe bølger
Under søvn eller under vågne tilstande, når et dyr hviler eller på anden måde ikke er engageret i sine omgivelser, viser hippocampus EEG et mønster af uregelmæssige langsomme bølger, der er noget større i amplitude end theta-bølger. Dette mønster afbrydes lejlighedsvis af store bølgeslag, de såkaldte skarpe bølger. Disse hændelser er forbundet med udbrud af spikeaktivitet, der varer 50-100 msek. i pyramidale celler i CA3 og CA1. De er også forbundet med kortvarige højfrekvente EEG-svingninger kaldet “ripples”. Ripples, med frekvenser i intervallet 150-200 Hz hos rotter, kan normalt kun påvises af elektroder, der enten er placeret inde i eller meget tæt på CA1-cellekropslaget. I modsætning hertil vil elektroder, der er placeret hvor som helst i hippocampus eller endda i tilstødende hjernestrukturer, ofte opfange skarpe bølger som store langsomme EEG-afbøjninger, der varer 200-400 msek.
I rotter er skarpe bølger mest robuste under søvn, hvor de forekommer med en gennemsnitlig frekvens på omkring 1 pr. sekund, men i et meget uregelmæssigt tidsmønster. Skarpe bølger forekommer også under inaktive vågenhedstilstande, men de er mindre hyppige og normalt mindre. Skarpe bølger er også blevet observeret i menneskets tindingelapper og i abernes hippocampus.I aber er skarpe bølger ret robuste, men de forekommer ikke nær så hyppigt som hos rotter.
Et af de mest interessante aspekter ved skarpe bølger er, at de synes at være forbundet med hukommelse. Wilson og McNaughton 1994 og talrige senere undersøgelser har rapporteret, at når hippocampale stedceller har overlappende rumlige fyringsfelter (og derfor ofte fyrer nær-simultanitet), har de tendens til at vise korreleret aktivitet under søvn efter den adfærdsmæssige session. Denne forstærkning af korrelationen, almindeligvis kendt som reaktivering, har vist sig at være begrænset til hovedsageligt til skarpe bølger. Det er blevet foreslået, at skarpe bølger i virkeligheden er reaktiveringer af neurale aktivitetsmønstre, der blev lagret i hukommelsen under adfærden, og som drives af en styrkelse af synaptiske forbindelser i hippocampus. Denne idé udgør en nøglekomponent i teorien om “to-trins-hukommelse”, som Buzsaki og andre er fortalere for, og som går ud fra, at minder lagres i hippocampus under adfærd og senere overføres til neocortex under søvn: skarpe bølger foreslås at drive hebbiske synaptiske ændringer i de neocortiske mål for hippocampus’ udgangsspor.
Evolution
Hippocampus har generelt samme udseende på tværs af pattedyrarter, fra basale arter som pindsvinet til de mest “avancerede” arter som mennesket.Hippocampus-størrelsen i forhold til kropsstørrelsen stiger generelt og er ca. dobbelt så stor hos primater som hos pindsvinet. Den stiger dog ikke i nærheden af den samme hastighed som forholdet mellem neocortex og kropsstørrelse.Hippocampus optager således et meget større volumen af den kortikale kappe hos gnavere end hos primater.
Der er også en generel sammenhæng mellem størrelsen af hippocampus og rumlig hukommelse: når der foretages sammenligninger mellem lignende arter, har de arter, der har en større kapacitet til rumlig hukommelse, tendens til at have større hippocampusvolumener. Dette forhold gælder også for kønsforskelle: hos arter, hvor hanner og hunner udviser store forskelle i evnen til rumlig hukommelse, er der også en tendens til, at de udviser tilsvarende forskelle i hippocampusvolumen
Non-mammalian arter har ikke en hjernestruktur, der ligner pattedyrenes hippocampus, men de har en, der anses for at være homolog til denne. Som påpeget ovenfor er hippocampus i det væsentlige den mediale kant af hjernebarken. Kun pattedyr har en fuldt udviklet cortex, men den struktur, som den er udviklet fra, kaldet pallium, findes hos alle hvirveldyr, selv de mest primitive som f.eks. lampret eller hagfisk. Pallium er normalt opdelt i tre zoner: medial, lateralt og dorsalt. Det mediale pallium danner forløberen for hippocampus. Det ligner ikke hippocampus visuelt, fordi lagene ikke er forvredet i en S-form eller omsluttet af gyrus dentata, men homologien er indikeret af stærke kemiske og funktionelle lighedspunkter. Der er nu beviser for, at disse hippocampalignende strukturer er involveret i rumlig kognition hos fugle, krybdyr og fisk.
Hos fugle er overensstemmelsen tilstrækkelig veletableret til, at de fleste anatomikere omtaler den mediale palliale zone som “avianhippocampus”. Talrige fuglearter har stærke rumlige færdigheder,især dem, der gemmer føde i en cache. Der er tegn på, at fugle, der gemmer mad, har en større hippocampus end andre fuglearter, og at skader på hippocampus forårsager forringelser af den rumlige hukommelse.
Historien for fisk er mere kompleks. Hos teleostfisk (som udgør langt størstedelen af de eksisterende arter) er forhjernen mærkeligt forvrænget i forhold til andre typer hvirveldyr. De fleste neuroanatomikere mener, at teleosternes forhjerner i det væsentlige er omvendte, som en sok vendt på vrangen, således at strukturer, der ligger indeni ved siden af hjertekamrene hos de fleste hvirveldyr, findes på ydersiden hos teleosterne og omvendt. En af konsekvenserne af dette er, at man mener, at det mediale pallium (“hippocampal” zone) hos et typisk hvirveldyr svarer til det laterale pallium hos en typisk fisk. Flere fisketyper (især guldfisk) har eksperimentelt vist sig at have en stærk rumlig hukommelse og endda at danne “kognitive kort” over de områder, de opholder sig i. Der er beviser for, at skader på lateral pallium forringer den rumlige hukommelse (navigation over store afstande, som f.eks. laksens hjemstavn, synes dog at være afhængig af andre mekanismer).
Hippocampusregionens rolle i navigationen synes således at begynde langt tilbage i hvirveldyrenes evolution og går forud for opdelinger, der fandt sted for flere hundrede millioner år siden. Det vides endnu ikke, om det mediale pallium spiller en lignende rolle hos endnu mere primitive hvirveldyr, såsom hajer og rokker eller endog lampretter og haglfisk. Nogle insektarter og bløddyr som blæksprutter har også stærke evner til rumlig indlæring og navigation, men disse synes at fungere anderledes end det rumlige system hos pattedyrene, så der er endnu ingen god grund til at tro, at de har en fælles evolutionær oprindelse; der er heller ikke tilstrækkelig stor lighed i hjernestrukturen til, at man kan identificere noget, der ligner en “hippocampus” hos disse arter.
Se også
- Amygdalohippocampectomy
- Median forebrain bundle
- Septal nuclei
Referencer & Bibliografi
Nøgletekster
Bøger
- Aboitiz, F, Morales D, Montiel J (2003). Den evolutionære oprindelse af pattedyrs isocortex: På vej mod en integreret udviklingsmæssig og funktionel tilgang. Behav. Brain Sciences 26: 535-552.
- Amaral, D; Lavenex P (2006). “Ch 3. Hippocampal Neuroanatomy” Andersen P, Morris R, Amaral D, Amaral D, Bliss T, O’Keefe J The Hippocampus Book, Oxford University Press.
- Broglio, C, Gómez A, Durán E, Ocaña FM, Jiménez-Moya F, Rodríguez F, Salas C (2002). Kendetegn ved en fælles plan for forhjernen hos hvirveldyr: Specialiserede palliale områder til rumlig, tidsmæssig og følelsesmæssig hukommelse hos actinopterygiske fisk. Brain Res. Bull. 57: 397-399.
- Buzsáki, G (2002). Theta-svingninger i hippocampus. Neuron 33: 325-340.
- Buzsáki, G (2006). Rhythms of the Brain.
- Cantero, JL, Atienza M, Stickgold R, Kahana MJ, Madsen JR, Kocsis B (2003). Søvnafhængige theta-svingninger i den menneskelige hippocampus og neocortex. J. Neurosci. 23: 10897-10903.
- Colombo, M, Broadbent N (2000). Er fuglehippocampus en funktionel homolog af pattedyrs hippocampus?. Neurosci. Biobehav. Rev. 24: 465-484.
- Eichnbaum, H; Cohen NJ (1993). Memory, Amnesia, and the Hippocampal System, MIT Press.
- Ekstrom, AD, Kahana MJ, Caplan JB, Fields TA, Isham EA, Newman EL, Fried I (2003). Cellulære netværk, der ligger til grund for menneskelig rumlig navigation. Nature 425: 184-188.
- Gray, JA; McNaughton N (2000). Neuropsykologi af angst: An Enquiry into the Functions of the Septo-Hippocampal System, Oxford University Press.
- Green, JD, Arduini AA (1954). Hippocampal elektrisk aktivitet i arousal. J. Neurophysiol. 17: 533-557.
- Jacobs, LF, Gaulin SJ, Sherry DF, Hoffman GE (1990). Evolution af rumlig kognition: kønsspecifikke mønstre af rumlig adfærd forudsiger hippocampal størrelse. PNAS 87: 6349-6352.
- Jacobs, LF (2003). Udviklingen af det kognitive kort. Brain Behav. Evol. 62: 128-139.
- Kirk, IJ, McNaughton N (1991). Supramammillær cellefyring og hippocampal rytmisk langsom aktivitet. Neuroreport 11: 723-725.
- Maguire, EA, Gadian DG, Johnsrude IS, Good CD, Ashburner J, Frackowiak RS, Frith CD (2000). Navigationsrelaterede strukturelle ændringer i hippocampi hos taxachauffører. PNAS 97: 4398-4403.
- McNaughton, BL, Battaglia FP, Jensen O, Moser EI, Moser MB (2006). Stiintegration og det neurale grundlag for det “kognitive kort”. Nat. Rev. Neurosci. 7: 663-678.
- Moser, EI, Kropf E, Moser M-B (2008). Stedceller, gitterceller og hjernens rumlige repræsentationssystem. Ann. Rev. Neurosci. 31.
- Nieuwenhuys, R (1982). En oversigt over organiseringen af hjernen hos Actinopterygian fisk. Am. Zool. 22: 287-310.
- O’Kane, G, Kensinger EA, Corkin S (2004). Bevis for semantisk indlæring i dyb amnesi: En undersøgelse med patient H.M.. Hippocampus 14: 417-425.
- O’Keefe, J; Nadel L (1978). The Hippocampus as a Cognitive Map, Oxford University Press.
- Portavella, M, Vargas JP, Torres B, Salas C (2002). Virkningerne af telencefaliske palliale læsioner på rumlig, tidsmæssig og følelsesmæssig læring hos guldfisk. Brain Res. Bull. 57: 397-399.
- Rodríguez, F, Lópeza JC, Vargasa JP, Broglioa C, Gómeza Y, Salas C (2002). Rumlig hukommelse og hippocampal pallium gennem hvirveldyrs evolution: indsigt fra krybdyr og teleostfisk. Brain Res. Bull. 57: 499-503.
- Sainsbury, RS, Heynen A, Montoya CP (1987). Adfærdsmæssige korrelater af hippocampal type 2 theta i rotte. Physiol. Behav. 39: 513-519.
- Scoville, WB, Milner B (1957). Tab af nylig hukommelse efter bilaterale hippocampale læsioner. J. Neurol. Neurosurg. Psych. 20: 11-21.
- Shettleworth, SJ (2003). Hukommelse og hippocampal specialisering i fødevarelagrende fugle: Challenges for Research on Comparative Cognition. Brain Behav. Evol. 62: 108-116.
- Skaggs, WE, McNaughton BL, Wilson MA, Barnes CA (1996). Thetafasepræcession i hippocampale neuronale populationer og kompression af tidsmæssige sekvenser. Hippocampus 6: 149-176.
- Squire, LR (1992). Hukommelse og hippocampus: en syntese fra resultater med rotter, aber og mennesker. Psych. Rev. 99: 195-231.
- Squire, LR; Schacter DL (2002). The Neuropsychology of Memory, Guilford Press.
- Vanderwolf, CH (1969). Hippocampal elektrisk aktivitet og frivillig bevægelse hos rotte. EEG & Clin. Neurophysiol. 26: 407-418.
- Vargas, JP, Bingman VP, Portavella M, López JC (2006). Telencephalon og geometrisk rum hos guldfisk. Eur. J. Neurosci. 24: 2870-2878.
- West, MJ (1990). Stereologiske undersøgelser af hippocampus: en sammenligning af de hippocampale underopdelinger af forskellige arter, herunder pindsvin, laboratorie gnavere, vilde mus og mænd… Prog. Brain Res. 83: 13-36.
Papers
- Amaral DG og Cowan WM. 1980. Subkortikale afferenter til den hippocampale formation hos aben. Journal of Comparative Neurology. Feb 15; 189(4):573-91.
- Duvernoy, H. (2005) The Human Hippocampus, 3rd ed. Berlin: Springer-Verlag.
Yderligere materiale
Papirer
- Google Scholar
- http://www.psycheducation.org/emotion/hippocampus.htm Store hjerneture!!!
- BrainMaps at UCDavis hippocampus
- Artificial Hippocampus
- BrainInfo at the University of Washington hier-164
- John O’Keefe & Lynn Nadel (1978) The Hippocampus as a Cognitive Map , Oxford University Press. Fuld tekst
Yderligere billeder
Føj et billede til dette galleri
Menneskehjerne: Limbisk system
Amygdala – Cingulate gyrus – Fornicate gyrus – Hippocampus – Hypothalamus – Mammillary body – Nucleus accumbens – Orbitofrontal cortex – Parahippocampal gyrus
|
v-d-e
Menneskelig hjerne: Forhjernen (cerebrum – cerebral cortex – cerebrale hemisfærer, grå substans) (TA A14.1.09.002-240, 301-320, GA 9.818-826) |
|||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Frontallappen |
|
||||||||||||||||
| Parietallappen |
|
Superolateral
|
Medial/inferior
|
Både
|
|||||||||||||
| Occipitallappen |
|
Superolateral
|
Medial/inferior
|
||||||||||||||
| Temporallappen |
|
Superolateral
|
Medial/inferior
|
||||||||||||||
| Interlobar sulci/fissurer |
|
Superolateral
|
Medial/inferior
|
||||||||||||||
| Limbic lobe |
|
Andre
|
|||||||||||||||
| Insulær lobe |
Lang gyrus af insula – Kort gyri af insula – Cirkulær sulcus af insula |
||||||||||||||||
| Generelt |
Operculum – Polerne af hjernehalvdelene |
||||||||||||||||
Nogle kategoriseringer er tilnærmelser, og nogle Brodmann-områder spænder over gyri.
|
|||||||||||||||||
Denne side bruger indhold med Creative Commons-licens fra Wikipedia (se forfattere).