- Introduction
- Metodit
- Jakaumatiedot ja biomiluokitus
- Lajikoostumus eri biomien välillä
- Ominaisuustiedot
- Ominaisuushypervolyymeja mittaamalla
- Toiminnallinen erottuvuus ja laajalle levinneisyys
- Funktionaalinen tila ja eliölajien samankaltaisuus
- Tulokset
- Keskustelu
- Tietopuutteet
- Yleisiä kuvioita biomeissa
- Biomien väliset vertailut
- Johtopäätökset
- Tietojen saatavuus
- Tekijöiden panos
- Rahoitus
- Conflict of Interest Statement
- Kiitokset
- Supplementary Material
Introduction
Ekologit ja biogeografit järjestävät maakasvien biologisen monimuotoisuuden ilmastollisesti määräytyviin biomeihin, joiden fysiologiaa luonnehtivat hallitsevien lajien kasvumuodot ja toiminnalliset ominaisuudet (Moncrieff et al., 2016). Biomi-käsitteellä onkin pitkä historia, joka juontaa juurensa Carl Ludwig Willdenowiin ja Alexander von Humboldtiin. Willdenow havaitsi, että samankaltaiset ilmastot tukevat samankaltaisia kasvillisuusmuotoja, ja Humboldt havaitsi laajan yhteyden kasvien levinneisyyden, fysiologian ja ympäristötekijöiden välillä. Kaiken kaikkiaan biomin käsite kuvastaa olettamusta, jonka mukaan samankaltaiset ympäristöpaineet valitsevat lajeja, joilla on samankaltaisia toiminnallisia ominaisuuksia, riippumatta niiden evoluutiohistoriasta. Samaan aikaan maapallon nykyiset biomit ovat jossain määrin fylogeneettisesti erillisiä, ja monet tai useimmat niille ominaisista lajeista ovat peräisin tietyistä sukulinjoista, joilla on keskeisiä sopeutumismahdollisuuksia ilmastovalinnan lisäksi myös muihin paineisiin, kuten tulipaloon tai megaherbivoreihin (Woodward et al., 2004; Pennington et al., 2006; Donoghue ja Edwards, 2014). Koska biomit edustavat laajamittaisia säännönmukaisuuksia maapallon kasvillisuudessa, biomien välisten toiminnallisten erojen ymmärtäminen on ratkaisevan tärkeää globaalin hiilenkierron ja maapallojärjestelmän toiminnan mallintamisen kannalta, mukaan lukien reaktiot ihmisen aiheuttamiin globaaleihin muutoksiin (Bonan ym, 2012; van Bodegom et al., 2014; Xia et al., 2015).
Viimeisten kahden vuosikymmenen aikana kasvien biologisen monimuotoisuuden funktionaalisen ulottuvuuden ymmärtäminen mantereiden mittakaavoissa on lisääntynyt nopeasti (Swenson et al., 2012; Lamanna et al., 2014; Šímová et al., 2018). Laajat tietojen synteesi- ja analyysitoimet ovat määritelleet rajallisen funktionaalisen ominaisuusavaruuden, joka sisältää kasvin kokonaiskoon, lehtien talouden ja hydrauliikan vaihtelun, joka tunnetaan funktionaalisten ominaisuuksien spektreinä (Wright et al., 2004; Díaz et al., 2016). Funktionaalisten ominaisuuksien spektrit määrittelevät fysiologisia ja ekologisia kompromisseja, jotka määrittävät kasvien elämäntapastrategioita (Westoby, 1998; Reich ym., 2003; Craine, 2009) ja niiden vaikutusta yhteisöjen kokoamiseen (Kraft ym., 2008), ekosysteemin toimintaan (Lavorel ja Garnier, 2002; Garnier ym., 2004; Kerkhoff ym., 2005; Cornwell ym., 2008) ja jopa evoluutionopeuteen (Smith ja Donoghue, 2008; Smith ja Beaulieu, 2009). Kasvien funktionaalisten ominaisuuksien spektrien yleisyys ja hyödyllisyys on nopeuttanut niiden sisällyttämistä kasvillisuuden globaalia jakautumista (van Bodegom et al., 2014), biogeokemiallista kiertoa (Bonan et al., 2012) ja ekosysteemipalveluja (Díaz et al., 2007; Cadotte et al., 2011; Lavorel et al., 2011) koskeviin malleihin. Kuitenkin vain harvoissa tutkimuksissa on tarkasteltu kattavia toiminnallisen monimuotoisuuden mittareita biomin mittakaavassa sekä hallitsevien että alisteisten lajien ja elämänmuotojen osalta.
On olemassa kaksi vastakkaista ennustekokonaisuutta siitä, miten toiminnallinen monimuotoisuus vaihtelee ilmastollisesti ja fysiognomisesti erilaisten biomien välillä. Yhtäältä, jos eliölajien biologinen monimuotoisuus heijastaa vaihtelua käytettävissä olevassa ekologisessa kapeikkotilassa, taksonomisesti vähemmän monimuotoisten eliölajien pitäisi edustaa pienempää, pitkälti sisäkkäistä osajoukkoa monimuotoisempien eliölajien valtaamasta toiminnallisesta tilasta. Toisaalta kasvien rakenteen ja toiminnan perustavanlaatuisten kompromissien globaalin luonteen ja samankaltaisten lajeissa samalla tavalla vaikuttavien valintapaineiden vuoksi erilaisissa ympäristöissä elävät lajiryhmät voivat itse asiassa jakaa samankaltaisia piirreavaruuden alueita (Reich et al., 2003; Wright et al., 2004; Díaz et al., 2016). Viimeaikaiset tutkimukset vahvistavat, että ilmaston ja toiminnallisen ja taksonomisen monimuotoisuuden (eli lajirikkauden) väliset suhteet ovat monimutkaisia ja mittakaavariippuvaisia. Joissakin tapauksissa funktionaalinen monimuotoisuus seuraa tiiviisti ilmastollisia gradientteja, jolloin funktionaalinen monimuotoisuus on vähäisempää vaihtelevammissa ja äärimmäisemmissä olosuhteissa (Swenson et al., 2012; de la Riva et al., 2018), ja funktionaalisen tilan määrä paikallisissa kokoelmissa laajenee taksonomisen rikkauden kasvaessa (Lamanna et al., 2014; Li et al., 2018). Alueellisen mittakaavan lajistoanalyysit viittaavat kuitenkin siihen, että suuriin lajirikkauden muutoksiin voi liittyä vähäisiä vaikutuksia toiminnalliseen monimuotoisuuteen (Lamanna et al., 2014; Šímová et al., 2015). Lisäksi taksonomisen ja funktionaalisen monimuotoisuuden reaktiot ilmastoon voivat vaihdella tärkeimpien kasvien kasvumuotojen välillä (Šímová et al., 2018), ja eri kasvumuotojen suhteellinen monimuotoisuus voi muuttua merkittävästi eri biomeja määrittelevien ilmastogradienttien varrella (Engemann et al., 2016).
Funktionaalisen monimuotoisuuden analysoinnissa mantereelta globaaliin mittakaavaan on edistytty osittain siksi, että biologisen monimuotoisuuden taksonomisia, levinneisyydeltään erilaisia, filogeneettisiä ja funktionaalisia aspekteja ei ole saatu riittävän kattavasti katalogisoitua tarpeeksi aineistoon perustuvia tietoja (Hortal et al., 2015). Lisäksi, vaikka tietoja on saatavilla rajoitetusti, lajien maantieteellisiä levinneisyyksiä, toiminnallisia piirteitä ja fylogeneettisiä suhteita kuvaavien suurten, toisistaan poikkeavien tietokokonaisuuksien standardointiin ja integrointiin liittyy merkittäviä tietoteknisiä haasteita (Violle ym., 2014). Tässä tutkimme kasvilajien funktionaalisen monimuotoisuuden jakautumista Pohjois- ja Etelä-Amerikan biomeissa Botanical Information and Ecology Network -tietokannan (BIEN; Enquist et al., 2016; Maitner et al., 2018) avulla, joka kokoaa yhteen levinneisyys- ja funktionaalisten ominaisuuksien tiedot >100 000 maakasvilajista. Erityisesti tarkastelemme biomien toiminnallisen monimuotoisuuden ja erottuvuuden vaihtelua, joka perustuu 100 × 100 km:n ruudukkosolujen mittakaavassa oleviin maakasvilajien levinneisyyskarttoihin ja kattavaan tietokokonaisuuteen kuudesta toiminnallisesta ominaisuudesta, jotka heijastavat ekologisten strategioiden vaihtelun pääakseleita.
Tutkimuksemme tavoitteet tässä tutkimuksessa ovat kolminkertaiset. Ensinnäkin, korostaaksemme pysyviä tietovajeita, dokumentoimme käytettävissä olevan tiedon laajuuden, joka luonnehtii biomien toiminnallista monimuotoisuutta ja erottuvuutta. Toiseksi, ottaen huomioon käytettävissä olevat tiedot, kuvaamme toiminnallisen monimuotoisuuden jakautumista biomien sisällä sekä hallitsevien että alisteisten kasvumuotojen välillä. Näiden analyysien avulla voimme paremmin kvantifioida biomin toiminnallisen erottuvuuden tunnistamalla biomin yleisimpien lajien yleisimmät toiminnalliset strategiat. Kolmanneksi kysymme, onko biomeille itse asiassa ominaista toiminnallisesti erilliset lajikokoelmat, jotka perustuvat moniulotteisten hypervolyymeiden päällekkäisyyteen toiminnallisten ominaisuuksien avaruudessa.
Metodit
Jakaumatiedot ja biomiluokitus
Vähentääksemme useista eri lähteistä kootuille esiintymisaineistoille ominaisten otantaharhojen vaikutuksia käytimme BIEN 2.0 -levinneisyysalueiden aluekarttoja, jotka koskivat 88 417:ää eri puolilla Pohjois- ja Etelä-Amerikkaa levittäytynyttä kasvilajia (Goldsmith et al., 2016). BIEN-tietokantaan on integroitu standardoituja kasvihavaintoja, jotka ovat peräisin herbaarionäytteistä ja kasvillisuuslohkojen inventoinneista. BIEN-tietokannoissa olevat lajien levinneisyysaluekartat arvioitiin käyttäen yhtä kolmesta lähestymistavasta sen mukaan, mitä esiintymiä kustakin lajista oli saatavilla. Niiden lajien osalta, joilla oli vain yksi tai kaksi esiintymää (n. 35 % lajeista), maantieteellinen levinneisyysalue määriteltiin kutakin datapistettä ympäröivänä neliönä 75 000 km2 :n alueena. Niiden lajien maantieteelliset alueet, joilla oli kolme tai neljä esiintymää, määritettiin kuperan rungon avulla (noin 15 % lajeista). Lopuksi niiden lajien levinneisyysaluekartat, joilla oli vähintään viisi esiintymää, saatiin käyttämällä Maxent-lajin levinneisyyden mallintamisalgoritmia, jossa käytettiin 19 ilmastokerrosta ennustemuuttujina ja 19 spatiaalista ominaisvektoria suodattimina, joilla rajoitettiin mallien ennusteita (ks. Goldsmith et al., 2016, jossa on lisätietoja levinneisyyskarttamenetelmästä).
Levitimme BIEN 2.0:n kasvilajikartat 100 × 100 km:n ruudukkokartalle Lambert Azimuthal Equal Area -projektiolla saadaksemme lajin esiintymis-/esiintymismatriisin kullekin ruudukkosolulle. Olsonin et al. (2001) biomiluokituksen perusteella määrittelimme jokaisen matriisin ruudukkosolun johonkin biomiluokkaan. Myöhempiä analyysejä varten tarvittavien laskennallisten rajoitusten vuoksi yhdistimme joitakin biomeja niiden ilmaston ja kasvillisuuden samankaltaisuuksien sekä kirjallisuuden perusteella, jotta saimme taulukossa 1 kuvatun laajan luokituksen (lukuun ottamatta sisävesiä, kalliota ja jäätä sekä mangroveja). Chacon ja Caatingan ekosysteemialueet luokiteltiin vastaavasti Xeric Woodlands ja Dry forest Pradon ja Gibbsin (1993), Pennington et al. (2000), Banda et al. (2016), Silva de Miranda et al. (2018) mukaan.
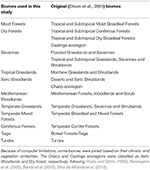
Taulukko 1. Taulukko 1. Chacon ekosysteemit. Yleiskatsaus tässä tutkimuksessa omaksuttuun biomiluokitukseen ja vastaavaan (Olson et al., 2001) biomiluokitukseen (pois lukien sisävedet, kallio ja jää sekä mangrovet).
Lajikoostumus eri biomien välillä
Ymmärtääksemme funktionaalisen ominaisuusavaruuden vaihtelua eri biomien välillä selvitimme ensin eroja kasvilajien lajikoostumuksessa biomien välillä. Kunkin ruudukkosolun ja biomin lajiluettelon perusteella määrittelimme kullekin biomille tyypillisiksi lajeiksi ne lajit, joiden suurin osa levinneisyysalueesta on kyseisessä biomissa. Näitä tietyn biomin maantieteellisesti vallitsevien lajien luetteloita verrattiin muiden biomien lajiluetteloihin. Tämä pareittainen vertailu tarjoaa yksinkertaisen tavan päätellä taksonomista päällekkäisyyttä biomien välillä (eli sitä, kuinka suuri osuus yhden biomin vallitsevista lajeista on yhteisiä toisen biomin kanssa). Niistä 88 417 lajista, joiden levinneisyysaluekartat ovat saatavilla, 44 899 lajin levinneisyysalue kattaa useamman kuin yhden biomin, ja 43 518 lajia on endeemisiä tietylle biomille.
Ominaisuustiedot
Purimme kaikki BIEN 3.0 -tietokannassa (haettu 7.2.2018) saatavilla olevat Uuden maailman kasvilajien ominaisuustiedot, jolloin saimme kaikkiaan 80 405 lajitason ominaisuushavaintoja. Tämän jälkeen suodatimme tiedot kuudesta toiminnallisesta ominaisuudesta: kasvin maksimikorkeus (m), siementen massa (mg), puun tiheys (mg/cm3), lehtien ominaispinta-ala SLA (cm2/g) ja lehtien fosfori- ja typpipitoisuus massayksikköä kohti (mg/g). Suodatuksen jälkeen jäljelle jäi yhteensä 18 192 lajitason havaintoa. Näistä 8 820 lajin osajoukkoa, jossa oli sekä levinneisyyskarttoja että ominaisuustietoja, käytettiin jatkoanalyyseihin.
Kunkin biomin funktionaalisen ominaisuusavaruuden arvioimiseksi tarvitsimme täydelliset ominaisuustiedot. Tästä syystä laskimme puuttuvat ominaisuustiedot fylogeneettisesti (Bruggeman ym., 2009; Penone ym., 2014; Swenson, 2014; Swenson ym., 2017) käyttäen R-pakettia “Rphylopars” v 0.2.9 (Goolsby ym., 2017) ja Smithin ja Brownin (2018) hiljattain julkaisemaa siemenkasvien fylogeniaa perustana. Fylogeneettinen imputaatio on työkalu puuttuvien tietojen ennustamiseen funktionaalisten ominaisuuksien tietokokonaisuuksissa, joka perustuu oletukseen, että läheisesti sukua olevilla lajeilla on yleensä samanlaiset ominaisuuksien arvot (Swenson, 2014). Käytimme ALLBM-puuta (eli geenipankki ja Open Tree of life -taksonit, joiden selkärangan on toimittanut Magallón et al., 2015), koska se maksimoi päällekkäisyyden niiden lajien välillä, joilla on käytettävissä olevia ominaisuus- ja levinneisyystietoja. Ominaisuusimputoinnin jälkeen jäljelle jäi yhteensä 7842 lajia, joilla oli täydelliset ominaisuustiedot ja levinneisyyskartat, jatkoanalyysejä varten.
Ominaisuushypervolyymeja mittaamalla
Määritimme biomien suhteellista toiminnallista monimuotoisuutta laskemalla ominaisuushypervolyymeja lajipoolin lajipooleista biomien solujen sisällä. Laskennallisten rajoitusten vuoksi muodostimme ominaisuuksien hypervolyymeja käyttäen satunnaisotoksen, joka koostui 20 prosentista kunkin biomin soluista. Kunkin ruudukkosolun hypervoluumit estimoitiin käyttämällä uutettuja kuutta toiminnallista ominaisuutta ja R-pakettia “hypervolume” (Blonder et al., 2014, 2018) käyttäen Gaussin KDE-menetelmää, jossa on oletuksena Silvermanin kaistanleveysestimaattori. Siemenmassa, korkeus ja puun tiheys muunnettiin log-muunnoksella, kun taas SLA muunnettiin neliöjuurimuunnoksella. Kaikki ominaisuudet skaalattiin ja keskitettiin ennen analyysia. Hypervolyymit ilmoitetaan standardipoikkeamien (sd) yksikköinä käytettyjen ominaisuuksien lukumäärän potenssiin (eli sd6).
Toiminnallinen erottuvuus ja laajalle levinneisyys
Koska biomeille on ominaista niiden vallitseva kasvillisuus, tarkastelimme myös lajien maantieteellistä yleisyyttä ja toiminnallista erottuvuutta biomien sisällä. Lajien levinneisyysaluekarttojen ja funktionaalisia piirteitä koskevien tietojen avulla arvioimme kunkin lajin funktionaalista erottuvuutta ja laajalle levinneisyyttä kussakin eliölajistossa Viollen ym. (2017) funktionaalisen harvinaisuuden käsitteellistä kehystä noudattaen. Käyttämällä koko (mitattujen ja laskennallisten) ominaisuuksien joukkoa mittasimme ensin kaikkien lajiparien välisen euklidisen etäisyyden standardoidussa ominaisuuksien avaruudessa. Tämän jälkeen laskimme kunkin lajin funktionaalisen erottuvuuden (Di) kussakin elinympäristössä lajin keskimääräisenä funktionaalisena etäisyytenä N muuhun lajiin elinympäristön lajipoolissa. Di skaalattiin 0:n ja 1:n välille kussakin elinympäristössä siten, että pienemmät arvot edustavat lajeja, jotka ovat toiminnallisesti yhteisiä (redundantteja), ja suuremmat arvot edustavat lajeja, jotka ovat toiminnallisesti erottuvia verrattuna muihin elinympäristön lajeihin. Arvioimme myös lajin maantieteellisen levinneisyyden (Wi) elinympäristössä, joka mitataan lajin elinympäristössä asuttamien ruudukkosolujen lukumääränä suhteessa kyseisen elinympäristön solujen kokonaislukumäärään. Arvo 1 tarkoittaa, että kyseinen laji esiintyy kaikissa biomin kattamissa ruudukkosoluissa. Sekä toiminnallinen erottuvuus että maantieteellinen levinneisyys laskettiin käyttämällä R-pakettia “funrar” (Grenié et al., 2017).
Koska meillä ei ole kattavia, paikalliseen runsauteen tai biomassaan perustuvia dominanssimittareita, käytimme mittareita Di ja Wi yksilöidäksemme kullekin eliölajistolle “yleisimmät” lajit lajeina, jotka ovat sekä maantieteellisesti levinneet laajalle (Wi > 0,5) että toiminnallisesti samankaltaisia lajeja (Di < 0,25). Viimeksi mainittua kynnysarvoa käytettiin, koska elinympäristöjen funktionaalisen erottuvuuden arvojen kolmas kvartiili vaihteli 0,2:sta 0,3:een. Erottaaksemme toiminnallisesti erilliset lajit toiminnallisesti samankaltaisista lajeista myöhemmissä analyyseissä käytimme siksi raja-arvona arvoa 0,25 (ts, lajeja, joiden erottuvuusarvot olivat < 0,25, pidettiin funktionaalisesti yhteisinä tai redundantteina tietyssä eliölajistossa).
Funktionaalinen tila ja eliölajien samankaltaisuus
Arvioidaksemme eliölajien hypervolyymeja koskevaa päällekkäisyyttä käytimme Sørensenin samankaltaisuusindeksiä käyttäen i) lajien kokonaismäärää ja ii) luetteloa lajeista, joita pidettiin funktionaalisesti yleisinä ja maantieteellisesti levinneinä kullekin eliölajistolle. Toiminnallisten ominaisuuksien tilojen samankaltaisuus laskettiin pareittaisena hypervolymien osittaisena päällekkäisyytenä biomien välillä. Murto-osittainen päällekkäisyys laskettiin jakamalla kahden hypervolyymin leikkauspisteen tilavuus kaksinkertaisesti niiden liittymien tilavuudella. Kaikki hypervoluumit arvioitiin käyttäen R-pakettia “hypervolume” (Blonder et al., 2014, 2018) käyttäen Gaussin KDE-menetelmää, jossa on oletuksena Silvermanin kaistanleveysestimaattori.
Tulokset
Havaitsimme huomattavaa päällekkäisyyttä kasvilajikoostumuksessa eri biomien välillä mallinnettujen lajien levinneisyysalueiden leikkauspisteiden perusteella (ks. kuva 1 ja lisätaulukko 1). Tämä taksonominen päällekkäisyys on suurempaa trooppisissa ja lauhkeissa biomeissa, ja näillä kahdella ilmastovyöhykkeellä on suhteellisen vähän yhteisiä lajeja. Mielenkiintoista on, että kuivilla metsäalueilla on yhteisiä lajeja sekä trooppisten että lauhkeiden biomien kanssa. Huolimatta trooppisille kosteille metsille ominaisten lajien suuresta osuudesta (83 %) tämä biomi jakaa suuren määrän lajeja myös muiden trooppisten biomien, kuten kuivien metsien, savannien, kuivien metsäalueiden ja trooppisten niittyjen kanssa. Lauhkeiden biomien, kuten lauhkeiden niittyjen, välimerellisten metsäalueiden, taigan ja tundran, suuri taksonominen päällekkäisyys viittaa mahdolliseen vähäiseen funktionaaliseen erillisyyteen, jolloin jotkin lauhkeat biomit edustavat lajirikkaampien ja funktionaalisesti monimuotoisempien trooppisten biomien heikompaa osajoukkoa.
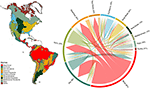
KUVIO 1. Lauhkeat biomit. Kasvilajien päällekkäisyys Uuden maailman biomeissa. Prosenttiarvot ilmaisevat niiden biomeissa esiintyvien lajien osuuden, joiden maantieteellisestä levinneisyysalueesta suurin osa on kyseisessä biomissa, N = 84 413. Kunkin haarautuneen nuolen pohja on sijoitettu siten, että se osoittaa sen biomin, joka sisältää suurimman osan lajin levinneisyysalueesta, kun taas leveys edustaa niiden lajien määrää, jotka ovat yhteisiä nuolen kärjessä olevien biomien kanssa. Taustalla oleva matriisi on lisätaulukossa 1.
Koska alle 10 prosenttia kartoitetuista lajeista oli edustettuna, ominaisuustiedot olivat melko niukkoja biomin mittakaavan lajikokoonpanojen osalta (kuva 2A). Lisäksi käytettävissä olevat ominaisuustiedot vaihtelivat huomattavasti ominaisuuksien, kasviklastien ja biomien välillä. Fylogeneettisesti tarkasteltuna lehtien P-arvo ja puun tiheys olivat erityisen heikosti edustettuina, ja kokonaisia klasteja ei ollut käytettävissä lainkaan tietoja. Tämän seurauksena noin 2/3 niistä 7 842 lajin ominaisuuksien arvoista, jotka oli laskettava, jouduttiin määrittämään siten, että monille niistä ei ollut läheistä sukulaistaksonia. Maantieteellisesti katsottuna SLA, pituus ja siemenmassa ovat alimitoitettuja tropiikissa ja lauhkeassa Etelä-Amerikassa, kun taas lehtien N-arvo ja puun tiheys ovat tasaisemmin edustettuina eri alueilla. Lehtien P:n näytteenotto on heikkoa kaikissa biomeissa. Ominaisuuksien hypervolyymeissä, jotka on luotu lajipooleista, jotka koostuvat 20 prosentin satunnaisvalikoimasta soluista kunkin biomin osalta, on lähinnä eroja trooppisten ja lauhkean/kylmän biomin välillä (kuva 2B). Ominaisuuksien hypervolyymeilla on taipumus olla suurempia ja vaihtelevampia trooppisissa biomeissa, ja vaihtelu on suurinta kosteissa ja kuivissa trooppisissa metsissä. Lauhkeista/kylmistä biomeista havupuuvaltaiset ja lauhkeat sekametsät tukevat suurimpia ominaisuuksien hypervolyymeja. Mielenkiintoista on, että kuivissa pensaikoissa hypervolymien jakauma muistuttaa enemmän trooppisia kuin lauhkeita biomeja.
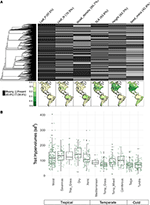
Kuva 2. (A) Niiden lajien osuus BIEN 3.0 -tietokannassa, joilla on tunnetut (harmaa) tai puuttuvat (musta) ominaisuusarvot. Vasemmalla oleva fylogenia vastaa Smithin ja Brownin (2018) siemenkasvien ALLBM-puuta (eli GenBankin ja Open Tree of life -taksoneita, joiden selkärangan on toimittanut Magallón et al., 2015). Kartat edustavat ominaisuustietoja sisältävien lajien osuutta suhteessa BIEN 2.0 -tietokannan lajien kokonaismäärään. (B) 20 %:n satunnaisesti valittujen 100 × 100 km:n solujen ominaisuuksien hypervolyymeiden jakautuminen kussakin eliölajissa. Hypervoluutiot ilmoitetaan standardipoikkeamien yksikköinä käytettyjen ominaisuuksien lukumäärän potenssiin.
Funktionaalisen erottuvuuden ja maantieteellisen levinneisyyden välinen suhde on huomattavan samankaltainen eri biomeissa (kuva 3). Jokaisella biomilla valtaosa lajeista on sekä maantieteellisesti rajoitettuja että toiminnallisesti samankaltaisia. Samalla jokaisen biomin toiminnallisesti erottuvimmat lajit olivat yleensä myös maantieteellisesti rajoittuneita, ja maantieteellisesti laajimmalle levinneet lajit olivat lähes aina toiminnallisesti samankaltaisia.
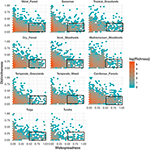
Kuva 3. Toiminnallisen erottuvuuden mallit eri biomeissa. Erottuvuus kuvaa sitä, miten lajit ovat toiminnallisesti etäällä toisistaan biomin sisällä (eli keskimääräinen pareittainen fenotyyppinen etäisyys polttoainelajin ja kaikkien muiden lajien välillä). Mitä suurempi arvo on, sitä kauempana laji on biomin funktionaalisen tilan keskipisteestä. Levinneisyys mittaa sitä, kuinka yleinen laji on maantieteellisesti elinympäristössä. Arvo 0 tarkoittaa, että laji esiintyy yhdessä ainoassa biomin solussa. Bin-arvot ovat lajien lukumäärä loge-asteikolla. Suorakulmiot edustavat lajeja, joita pidetään toiminnallisesti samanlaisina ja laajalle levinneinä kussakin biomissa käyttäen erottuvuuden raja-arvoja 0,25 ja levinneisyyden raja-arvoja 0,5.
Havaitsimme huomattavaa vaihtelua kasvumuotojen suhteellisessa jakautumisessa biomien sisällä ja niiden välillä. Puulajit edustavat hallitsevia kasvumuotoja trooppisissa biomeissa lajimäärissä mitattuna, kun taas ruohovartiset lajit hallitsevat lauhkeissa ympäristöissä (kuva 4). Kun otimme huomioon vain ne lajit, jotka ovat laajalle levinneitä ja toiminnallisesti yleisiä kussakin elinympäristössä, kasvumuotojen jakautuminen eri elinympäristöissä muuttui, erityisesti puiden, yrttien ja ruohojen osuuden osalta. Esimerkiksi trooppisissa biomeissa puiden osuus väheni kaikissa biomeissa kosteaa metsää lukuun ottamatta. Lauhkeissa biomeissa ruohojen osuus kasvoi erityisesti napa-alueilla. Mielenkiintoista on, että kasvumuotojen jakauma kserisissä metsissä muistuttaa enemmän jakaumaa lauhkeissa biomeissa, kun tarkastellaan vain laajalle levinneitä ja toiminnallisesti yleisiä lajeja.
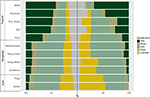
Kuva 4. Kasvumuotojen jakautuminen kullakin biomilla (vasemmalla) käyttämällä lajien kokonaismäärää ja (oikealla) käyttämällä vain niitä lajeja, jotka ovat toiminnallisesti samankaltaisia ja laajalle levinneitä biomissa (ks. kuva 3).
Lajikoostumuksen parittaiset vertailut eri biomien välillä paljastavat kolme pääklusteria, jotka edustavat trooppista, lauhkeaa ja polaarista ilmastovyöhykettä (kuva 5A) ja jotka heijastavat taksonomisia päällekkäisyyksiä suuressa määrin näiden alueiden sisällä, mutta eivät niiden välillä (kuva 1). Ominaisuuksien hypervolyymeja koskevat parittaiset vertailut biomien välillä osoittavat, että ilmastovyöhykkeet eivät ole yhtä selviä klustereita (kuva 5B). Tässä tapauksessa kseristen metsäalueiden toiminnallinen tila on merkittävästi päällekkäinen lauhkeiden biomien kanssa. Tämä analyysi paljastaa myös taigojen toiminnallisen tilan päällekkäisyyden lauhkean laidunmaan ja sekametsien kanssa. Ominaisuuksien hypervolyymeja pareittain vertailtaessa biomeja keskenään käyttäen vain niitä lajeja, joita pidetään funktionaalisesti yleisinä ja laajalle levinneinä, havaitaan vähemmän päällekkäisyyttä ominaisuuksien tiloissa ilmastovyöhykkeiden välillä ja niiden sisällä (kuva 5C). Vaikka kseriset metsät on nyt ryhmitelty muiden trooppisten biomien kanssa, näillä luontotyypeillä sekä trooppisilla ruohoalueilla on kuitenkin suurta päällekkäisyyttä toiminnallisessa tilassa lauhkean alueen biomien, kuten Välimeren alueen metsäalueiden ja lauhkean alueen ruohoalueiden, kanssa.
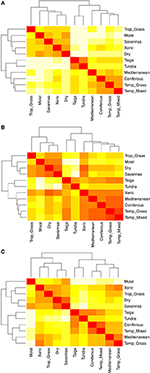
KUVIO 5. Elinympäristöt, joilla on suuri päällekkäisyys toiminnallisessa tilassa. (A) Lajikoostumuksen pareittainen erilaisuus biomien välillä. (B) Pareittainen eroavaisuus ominaisuuksien hypervolyymeissä (1-Sørensenin samankaltaisuus) biomien välillä käyttäen kokonaislajimäärää. (C) Pareittainen eroavuus ominaisuuksien hypervolyymeissa (1-Sørensenin samankaltaisuus) biomien välillä käyttäen ainoastaan niitä lajeja, joita pidetään toiminnallisesti samankaltaisina ja laajalle levinneinä. Mitä vaaleampi solu, sitä suurempi eroavaisuus.
Keskustelu
Analyysimme tuottavat kolme tärkeää oivallusta, jotka auttavat ymmärtämään maanpäällisiä eliölajeja funktionaalisen linssin kautta. Ensinnäkin osoitamme, että huolimatta edistymisestä ensisijaisten biodiversiteettitietojen kokoamisessa ja synteesissä on edelleen merkittäviä tietovajeita, jotka saattavat rajoittaa kykyämme kvantifioida biomien toiminnallista biodiversiteettiä mantereelta maailmanlaajuiseen mittakaavaan. Toiseksi käytettävissä olevien tietojen analyysimme osoittaa kuitenkin, että kaikilla Pohjois- ja Etelä-Amerikan biomeilla on huomattava yhteinen kuvio, jonka mukaan minkä tahansa biomin maantieteellisesti laajimmalle levinneillä lajeilla on taipumus jakaa yhteisiä funktionaalisia piirteitä, kun taas funktionaalisesti omaleimaisimpien lajien levinneisyys on poikkeuksetta rajoitettu. Kolmanneksi, kun tarkastellaan vain laajalle levinneitä ja toiminnallisesti yleisiä lajeja, biomit voidaan erottaa toiminnallisesti helpommin toisistaan, ja biomien väliset eroavaisuudet näyttävät heijastavan ilmaston ja kasvien toiminnallisen kapeikon välistä vastaavuutta. Kaiken kaikkiaan tuloksemme viittaavat siihen, että vaikka biomien funktionaalisen monimuotoisuuden tutkimus on vielä alkuvaiheessa, alan jatkokehitys tuottaa todennäköisesti oivalluksia, jotka yhdistävät evoluution, biogeografian, yhteisöjen kokoonpanon ja ekosysteemien toiminnan.”
Tietopuutteet
BIEN-tietokanta on erityisesti suunniteltu paikkaamaan aukkoja tietämyksessämme kasvien biologisesta monimuotoisuudesta, mutta, kuten Hortal ym. (2015) huomauttavatkin, keskenään vuorovaikutuksessa olevat tietopuutteet johtavat epävarmuuteen biologisen monimuotoisuuden määrittelemisessä suurimmissa mittakaavoissa. Esimerkiksi alle 10 prosentilla kartoittamistamme lajeista oli saatavilla sekä ominaisuustietoja että fylogeneettistä tietoa, ja lisäksi nämä puuttuvat ominaisuustiedot (~66 prosenttia lajien ominaisuustiedoista) olivat varsin jäsenneltyjä sekä fylogeneettisesti että maantieteellisesti. Näin ollen Raunkiaerin puute toiminnallisista ominaisuustiedoista (sensu Hortal et al., 2015) on vuorovaikutuksessa Wallacen puutteen kanssa levinneisyystiedoissa ja Darwinin puutteen kanssa fylogeneettisessä ymmärryksessä. Ja vaikka voimme käyttää fylogeneettistä tietoa näiden puuttuvien arvojen kohtuulliseen korvaamiseen (Swenson ym., 2017), useat tuloksena syntyvien kuvioiden näkökohdat, jotka ovat tärkeitä eliölajien luokittelun kannalta, ovat edelleen epävarmoja. Esimerkiksi olemassa olevat kasvumuototiedot viittaavat siihen, että puumaiset lajit hallitsevat kaikissa trooppisissa biomeissa, kun taas ruohomaisten kasvumuotojen suhteellinen monimuotoisuus on paljon suurempi lauhkeissa ja polaarisissa biomeissa (ks. myös Engemann ym., 2016; Zanne ym., 2018). Kuvastaako tämä leveysasteiden mukainen ruohomaisten kasvien suhteellisen monimuotoisuuden lisääntyminen näytteenottoharhoja ja trooppisten ruohomaisten kasvien taksonomisen tietämyksen puutetta vai heijastaako se erilaista evoluutio- ja biogeografista historiaa Nearctic- ja Neotropic-maailmoissa? Vallitsevat kasvumuodot ovat yksi tärkeimmistä biomeja erottavista piirteistä, joten systemaattiset muutokset monimuotoisimmissa kasvumuodoissa (olivatpa ne vallitsevia tai eivät) vaikuttavat väistämättä ennusteisiimme siitä, miten funktionaalinen monimuotoisuus saattaa muuttua eri biomeissa ja miten nämä muutokset vaikuttavat ekosysteemien toimintaan ja palveluihin.
Käytimme lajikarttoja syöttötietoina pääasiassa siksi, että pystyimme ratkaisemaan otantavirheiden kiistämättömän ongelman, joka on luonteenomaista useista eri lähteistä kootuille tietokokonaisuuksille (ks. täydentävä kuva 1). Lajien levinneisyysmallien perusteella laaditut levinneisyysaluekartat voivat kuitenkin olla epävarmuutta lisäävä tekijä tuloksissamme. Esimerkiksi ottaen huomioon tämän tutkimuksen maantieteellinen resoluutio ja tiettyjen biomien alueellinen monimutkaisuus, nämä mallit ovat voineet yliarvioida joidenkin lajien maantieteellisen laajuuden yhden biomin soluista läheisten ja toisiinsa liittyvien biomien soluihin. Esiintymistietojen käyttäminen ei muuttanut tämän tutkimuksen yleisiä tuloksia (ks. täydentävät kuvat 2-6); biomien välisen toiminnallisen päällekkäisyyden vähäinen väheneminen voisi kuitenkin paljastaa joidenkin lajien levinneisyysalueiden yliarvioinnin (ks. täydentävät kuvat 2, 5 ja 6). Esiintymistietojen käyttäminen muutti myös toiminnallisen erottuvuuden ja levinneisyyden välisiä yleisiä suhteita tuloksissamme, mikä johtui lauhkeiden ja trooppisten alueiden suhteellisesta ylinäytteenotosta (ks. täydentävä kuva 4). Tässä tapauksessa laajalle levinneisyyden mittariamme rajoittaa useimpien lajien erittäin rajallinen näytteenotto trooppisissa biomeissa. Kun tietokokonaisuudet ja näytteenotto paranevat, näitä epävarmuustekijöitä voidaan todennäköisesti vähentää tulevaisuudessa.
Toinen tietovaje, joka on korjattava, jotta biomien toiminnallista monimuotoisuutta voidaan ymmärtää, on runsaustietojen Prestonian puute. Runsaus on erityisen tärkeää toiminnallista monimuotoisuutta koskevissa tutkimuksissa, koska yhteisön kokoonpanon, ekosysteemin toiminnan ja biogeokemiallisen kierron kannalta tärkeimpiä ovat hallitsevimpien lajien ominaisuudet. Koska perustimme kokoelmamme lajien levinneisyysmalleista johdettuihin levinneisyyskarttoihin, pystyimme käsittelemään vain yleisyyden maantieteellistä komponenttia, mutta paikallisen runsauden ja maantieteellisen levinneisyyden välinen positiivinen suhde, niin sanottu miehityksen ja runsauden suhde, antaa viitteitä siitä, että kussakin elinympäristössä laajimmalle levinneet lajit saattavat usein olla myös runsaimpien joukossa (Borregaard ja Rahbek, 2010; Novosolov ym., 2017). Koska biomeja luonnehtivat yleensä alueella vallitsevat kasvumuodot, runsaustietojen integroiminen saattaa johtaa siihen, että biomeissa on vähemmän toiminnallista päällekkäisyyttä. Vaikka BIEN on koonnut runsaasti tonttitietoja, jotka mahdollistavat paikallisen runsauden arvioinnin, kohtuullisten vertailujen tekemistä eri alueiden ja kasvumuotojen välillä vaikeuttavat kuitenkin epätasainen näytteenotto, alueelliset erot gammadiversiteetissä ja epäyhtenäiset runsauden kvantifiointimenetelmät.
Yleisiä kuvioita biomeissa
Kannattavissa olevissa tiedoissa olevista jatkuvista aukoista huolimatta analyysimme paljastivat aiemmin dokumentoimattoman suhteen, joka on yhteinen kaikille läntisen pallonpuoliskon biomeille: missä tahansa biomissa laajimmalle levinneillä lajeilla on myös taipumus osoittaa vähäistä funktionaalista erottautuvuutta, kun taas funktionaalisesti erottautuvimmilla lajeilla on melkein poikkeuksetta rajoitettu levinneisyys (kuva 3). Tämä “miehitys-redundanssisuhde” voi viitata siihen, että tietyssä elinympäristössä vallitsevat ilmasto-olosuhteet valitsevat joukon yhteisiä ominaisuuksia, ja toiminnallisesti erottuvammat lajit rajoittuvat harvinaisempaan osajoukkoon elinympäristöjä kyseisessä elinympäristössä. Toiminnallisesti samankaltaisten lajien yleinen yleisyys kaikissa biomeissa ja “miehitys-redundanssi”-suhde ovat sopusoinnussa tuoreen maailmanlaajuisen analyysin kanssa yhteisötason toiminnallisesta monimuotoisuudesta, joka viittaa siihen, että elinympäristöjen suodattaminen johtaa toiminnallisesti samankaltaisten lajien rinnakkaiseloon (Li ym., 2018), sekä tutkimuksiin, jotka osoittavat, että toiminnalliset redundanssit lisäävät yhteisöjen vakautta (esim. Walker ym., 1999; Pillar ym., 2013). Lisäksi yhdessä sekä taksonomisen että funktionaalisen päällekkäisyyden suuren asteen kanssa biomien välillä (kuvat 1, 5) se, että yleiset, laajalle levinneet lajit ovat funktionaalisesti samankaltaisia, vahvistaa käsitystä siitä, että maakasveilla monenlaisissa ympäristöolosuhteissa on yhteisiä ominaisuuksia lähellä funktionaalisten ominaisuuksien spektrin ydintä (Wright et al., 2004; Díaz et al., 2016). Toisin kuin Umaña ym. (2017), meidän tuloksemme osoittavat, että harvinaiset, maantieteellisesti rajoitetut lajit voivat olla toiminnallisesti erilaisia kuin laajemmin levinneet ja yleiset lajit. Näin ollen avoimeksi jää kysymys siitä, ovatko toiminnallisesti erottuvammat lajit spesialisteja tietyissä ympäristöissä vai johtavatko niiden ominaisuusyhdistelmät demografisiin tai fysiologisiin kompromisseihin, jotka rajoittavat niiden maantieteellistä levinneisyyttä.
Biomien väliset vertailut
Vertailumme ominaisuuksien hypervolyymijakaumista eri biomien välillä (kuva 2B) on johdonmukainen sen havainnon kanssa, jonka mukaan lajistoltaan rikkaammat ympäristöt ovat samalla myös toiminnallisesti monimuotoisempia (Swenson et al., 2012; Lamanna et al., 2014; Li et al., 2018; Šímová et al., 2018). Tuloksemme ovat epäselvempiä sen hypoteesin suhteen, että kausiluonteiset ja äärimmäiset ilmastoympäristöt rajoittavat johdonmukaisesti lajien toiminnallista monimuotoisuutta (de la Riva ym., 2018). Kaikilla trooppisilla biomeilla on suurempi keskimääräinen toiminnallinen monimuotoisuus kuin kaikilla lauhkeilla biomeilla, ja polaarisilla biomeilla on todellakin pienimmät hypervolyymit. Kunkin ryhmän sisällä kuivemmilla tai kausivaihtelevammilla biomeilla ei kuitenkaan aina näy pienempiä hypervolyymeja (esim. kuivat metsät, kseriset metsäalueet), kuten olisimme odottaneet Currien ym. (2004) toleranssihypoteesin perusteella, kun taas optimaaliset ilmasto-olosuhteet tukevat useampia fysiologisten parametrien yhdistelmiä. Pelkästään näiden tulosten perusteella emme voi päätellä, ovatko lauhkeat ja polaariset biomit taksonomisesti vähemmän monimuotoisia toiminnallisen kapeikkotilan rajoitusten vuoksi vai ovatko niiden toiminnalliset hypervolyymit pieniä, koska ne eivät ole taksonomisesti monimuotoisia. Nollamallinnuslähestymistavat voisivat mahdollisesti auttaa erottamaan taksonomisen ja funktionaalisen monimuotoisuuden (Swenson ym., 2012; Lamanna ym., 2014; Šímová ym., 2018), mutta tällainen analyysi ei kuulunut tämän tutkimuksen piiriin. Tärkeämpää on, että tuloksemme vahvistavat sen ymmärtämisen tärkeyttä, miten evoluutio- ja biogeografinen historia muokkaavat eliölajien toiminnallista monimuotoisuutta (Woodward et al., 2004; Pennington et al., 2006; Donoghue ja Edwards, 2014; Moncrieff et al., 2016). Lajien (ja korkeampien sukulinjojen) laaja jakaminen eri biomeissa eri biogeografisten alueiden sisällä, mutta suurelta osin ei eri biogeografisten alueiden välillä, voisi sekä homogenisoida funktionaalista monimuotoisuutta alueiden sisällä että antaa vihjeitä ominaispiirteistä, joita erilaiset ympäristöt valitsevat tai vastustavat (Douma et al., 2012; Zanne ym., 2014, 2018).
Huolimatta huomattavasta hypervolyymin päällekkäisyydestä kaikkien biomien välillä (täydentävä kuva 2), trooppiset, lauhkeat ja kylmät biomit näyttävät kaikki vievän toisistaan erottuvia alueita toiminnallisessa tilassa (kuvat 5B,C). Tärkeimmät biomeja erottavat piirteet näyttävät olevan kasvien kokonaiskokoon liittyviä piirteitä, mukaan lukien sekä kypsä pituus että siementen massa (täydentävät kuvat 7-10), eivätkä niinkään lehtien taloudellisuuteen liittyviä piirteitä, kuten havaittiin paikallisemmissa, lohkoihin perustuvissa analyyseissä (esim. Douma et al., 2012). Poikkeuksena on lehtien P-pitoisuus, jossa oli huomattavia eroja trooppisten ja lauhkeiden/polaaristen biomien välillä (täydentävät kuvat 7-10), mikä on havaittu myös muissa analyyseissä, jotka perustuvat sekä lajikohtaisiin arvoihin että koko ekosysteemin mittauksiin (Kerkhoff ym., 2005; Swenson ym., 2012). Koska lehtien P-pitoisuus oli kaikkein harvimmin poimittu ominaisuus, trooppisten ja lauhkeiden/polaaristen alueiden väliset erot saattavat olla alttiita imputointimenettelyn aiheuttamille vääristymille. Lehtien P:llä on kuitenkin merkittävä fylogeneettinen signaali (Kerkhoff ym., 2006), mikä viittaa siihen, että imputaatioiden pitäisi olla harhattomia (Swenson ym., 2017). Leaf P on myös merkittävästi korkeampi ruohovartisissa kasvumuodoissa (Kerkhoff ym., 2006), joten leveyssuuntainen siirtymä pääasiassa puumaisesta monimuotoisuudesta ruohovartiseen monimuotoisuuteen (kuva 4) voi myös vaikuttaa tähän kuvioon.
Kun rajasimme hypervolyysianalyysit vain laajimmalle levinneisiin ja funktionaalisesti tavallisimpiin lajeihin kussakin eliölajistossa, yksittäiset eliölajit kussakin eliömaantieteellisessä valtakunnassa limittyivät funktionaalisen ominaisuuden tilassa vähemmän toistensa kanssa (vrt. kuviot 5B,C), mikä viittaisi siihen, että nämä lajit saattavat heijastella tiettyihin ympäristötekijöihin paremmin sopeutuneita fenotyyppejä. Tässä yhteydessä kserinen metsäbiomi on erityisen mielenkiintoinen. Osittain Chacon ekologisen alueen sisällyttämisen vuoksi kseriset metsäalueet ryhmittyvät taksonomisesti trooppisten biomien kanssa (kuva 5A). Mutta kun kaikkien lajien toiminnallisia hypervolyymeja tarkastellaan, ne ovat paljon samankaltaisempia kuin lauhkean vyöhykkeen biomit (kuva 5B). Kun lopuksi analysoidaan vain laajimmalle levinneitä ja toiminnallisesti yleisimpiä lajeja, kuivien metsäalueiden samankaltaisuus trooppisten biomien kanssa on jälleen lisääntynyt, mutta myös niiden samankaltaisuus lauhkeiden niittyjen ja Välimeren biomien kanssa säilyy korkeana (kuva 5C). Siirtymiä lämpimistä, mesisistä ympäristöistä kylmempiin, kuivempiin ja kausiluontoisempiin ympäristöihin helpottavat samankaltaiset piirteet, esimerkiksi ruohovartinen kasvutapa (Douma ym., 2012; Zanne ym., 2014, 2018), ja kylmempien ympäristöjen tapaan myös kuivien ympäristöjen yhteisöillä on taipumus olla fylogeneettisesti klusteroituneempia (Qian ja Sandel, 2017). Lisäksi kseristen vyöhykkeiden sijainti trooppisten alueiden välisen konvergenssivyöhykkeen rajalla tekee niistä maantieteellisen siirtymän trooppisen ja lauhkean alueen välillä. Se, että kseriset metsäalueet ovat sekä toiminnallisesti että biogeografisesti trooppisen ja lauhkean alueen välissä, vahvistaa edelleen ajatusta siitä, että ymmärtääksemme paremmin biomien toiminnallista monimuotoisuutta meidän on otettava huomioon niiden biogeografinen ja fylogeneettinen historia (Pennington ym., 2006; Donoghue ja Edwards, 2014; Moncrieff ym, 2016).
Johtopäätökset
Mikä tahansa maanpäällisten biomien luokittelu pakottaa pienen määrän erillisiä luokkia ilmastossa ja lajien levinneisyydessä esiintyviin jatkuviin gradientteihin ja edustaa siten monimutkaisen ekologisen maiseman karkeaa yksinkertaistamista (Moncrieff ym., 2016, mutta ks. Silva de Miranda ym., 2018). Huolimatta mahdollisesta liiallisesta yksinkertaistamisesta biomit ovat kuitenkin hyödyllisiä konstruktioita maapallon tärkeimpien maaekosysteemien biologisen monimuotoisuuden ja toiminnan järjestämisessä ja ymmärtämisessä, ja ominaisuuksiin perustuvilla lähestymistavoilla on suuri potentiaali auttaa mallintamaan dynaamisesti globaalia kasvillisuuden levinneisyyttä.
Tässä tutkimuksessa olemme osoittaneet, että suuri osa kaikkien biomien taksonomisesta moninaisuudesta edustaa lajeja, jotka ovat sekä kapeasti levinneitä että toiminnallisesti samanlaisia. Lisäksi kunkin biomin sisällä toiminnallisesti erottuvimmat lajit ovat yleensä myös maantieteellisesti harvinaisia, kun taas laajalle levinneillä lajeilla on tasaisesti alhainen toiminnallinen erottuvuus. Huolimatta biomien laajasta taksonomisesta ja toiminnallisesta päällekkäisyydestä ne ryhmittyvät erottuviksi, biogeografisesti ja ilmastollisesti erillisiksi yksiköiksi, erityisesti kun toiminnallinen ryhmittely perustuu kunkin biomin yleisimpiin ja toiminnallisesti yleisimpiin lajeihin. Biomien funktionaalisen ymmärryksen edistäminen edellyttää kuitenkin kasvillisuustyyppien sisäisen ja kasvillisuustyyppien välisen ominaisuusvaihtelun paremman luonnehdinnan lisäksi myös tietoa lajikokoonpanojen biogeografisesta ja fylogeneettisestä historiasta sekä lajien suhteellisesta runsaudesta biomien sisällä.
Tietojen saatavuus
Tässä tutkimuksessa analysoidut ominaisuustietokokonaisuudet ja lajien levinneisyysaluekartat ovat ladattavissa R-paketti BIEN:n välityksellä (Maitner et al., 2018). Katso http://bien.nceas.ucsb.edu/bien/ lisätietoja BIEN-tietokannasta.
Tekijöiden panos
SE-L, AK, BJE, DMN ja CV suunnittelivat tutkimuksen; SE-L ja AK analysoivat aineiston; SE-L ja AK kirjoittivat käsikirjoituksen ensimmäisen version; CV, BB, NK, BSM, BM, RKP, BS, SS, J-CS ja SKW osallistuivat piirteiden ja lajikarttatietokannan kokoamiseen ja luomiseen. Kaikki kirjoittajat osallistuivat käsikirjoituksen kehittämiseen ja kirjoittamiseen.
Rahoitus
SE-L, DMN ja AK saivat tukea Yhdysvaltain kansallisen tiedesäätiön myöntämästä tutkimusapurahasta (DEB-1556651). BJE:tä tukivat National Science Foundationin myöntämät palkinnot DEB-1457812 ja Macrosystems-1065861. CV sai tukea Euroopan tutkimusneuvostolta (apuraha ERC-StG-2014-639706-CONSTRAINTS) ja Ranskan biologisen monimuotoisuuden tutkimussäätiöltä (FRB; www.fondationbiodiversite.fr) CESAB-hankkeessa Causes and consequences of functionalararity from local to global scale. J-CS katsoo, että tämä työ on osa hänen VILLUM Investigator -hankettaan Biodiversity Dynamics in a Changing World, jota rahoittaa VILLUM FONDEN (apuraha 16549). SKW:tä tuki Uuden-Seelannin kauppa-, innovaatio- ja työministeriön tiede- ja innovaatioryhmän Strategic Science Investment Fund.
Conflict of Interest Statement
Tekijät ilmoittavat, että tutkimus tehtiin ilman kaupallisia tai taloudellisia suhteita, jotka voitaisiin tulkita mahdolliseksi eturistiriidaksi.
Kiitokset
Kiitämme Dan Griffithiä ja kahta arvioijaa rakentavasta kritiikistä, joka paransi tätä käsikirjoitusta huomattavasti. Kiitämme myös kaikkia BIEN-tiedonantajia (katso http://bien.nceas.ucsb.edu/bien/data-contributors/ täydellistä luetteloa varten).
Supplementary Material
Tämän artikkelin lisäaineisto löytyy verkosta osoitteesta: https://www.frontiersin.org/articles/10.3389/fevo.2018.00219/full#supplementary-material
Banda, R.K., Delgado-Salinas, A., Dexter, K. G., Linares-Palomino, R., Oliveira-Filho, A., Prado, D., et al. (2016). Kasvien monimuotoisuusmallit neotrooppisissa kuivissa metsissä ja niiden vaikutukset suojeluun. Science 353, 1383-1387. doi: 10.1126/science.aaf5080
PubMed Abstract | CrossRef Full Text | Google Scholar
Blonder, B., Lamanna, C., Violle, C. ja Enquist, B. J. (2014). N-ulotteinen hypervoluutio. Glob. Ecol. Biogeogr. 23, 595-609. doi: 10.1111/geb.12146
CrossRef Full Text | Google Scholar
Blonder, B., Morrow, C. B., Maitner, B., Harris, D. J., Lamanna, C., Violle, C., et al. (2018). Uusia lähestymistapoja n-ulotteisten hypervoluutioiden rajaamiseen. Methods Ecol. Evol. 9, 305-319. doi: 10.1111/2041-210X.12865
CrossRef Full Text | Google Scholar
Bonan, G. B., Oleson, K. W., Fisher, R. A., Lasslop, G., and Reichstein, M. (2012). Lehtien fysiologisten ominaisuuksien ja latvustovirtatietojen yhteensovittaminen: TRY- ja FLUXNET-tietokantojen käyttö yhteisön maamallin versiossa 4. J. Geophys. Res. Biogeosci. 117:G02026. doi: 10.1029/2011JG001913
CrossRef Full Text | Google Scholar
Borregaard, M. K., and Rahbek, C. (2010). Maantieteellisen levinneisyyden ja lajien runsauden välisen suhteen kausaalisuus. Q. Rev. Biol. 85, 3-25. doi: 10.1086/650265
PubMed Abstract | CrossRef Full Text | Google Scholar
Bruggeman, J., Heringa, J., ja Brandt, B. W. (2009). PhyloPars: puuttuvien parametriarvojen estimointi fylogenian avulla. Nucleic Acids Res. 37, W179-W184. doi: 10.1093/nar/gkp370
PubMed Abstract | CrossRef Full Text | Google Scholar
Cadotte, M. W., Carscadden, K. ja Mirotchnick, N. (2011). Beyond species: functional diversity and the maintenance of ecological processes and services. J. Appl. Ecol. 48, 1079-1087. doi: 10.1111/j.1365-2664.2011.02048.x
CrossRef Full Text | Google Scholar
Cornwell, W. K., Cornelissen, J. H., Amatangelo, K., Dorrepaal, E., Eviner, V. T., Godoy, O., et al. (2008). Kasvilajien ominaispiirteet vaikuttavat eniten karikkeen hajoamisnopeuteen eri puolilla maailmaa. Ecol. Lett. 11, 1065-1071. doi: 10.1111/j.1461-0248.2008.01219.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Craine, J. M. (2009). Luonnonvaraisten kasvien resurssistrategiat. New Jersey, NJ: Princeton University Press. doi: 10.1515/9781400830640
CrossRef Full Text | Google Scholar
Currie, D. J., Mittelbach, G. G., Cornell, H. V., Field, R., Guégan, J. F., Hawkins, B. A., et al. (2004). Ennusteet ja testit ilmastoon perustuvista hypoteeseista, jotka koskevat taksonomisen rikkauden laajamittaista vaihtelua. Ecol. Lett. 12, 1121-1134. doi: 10.1111/j.1461-0248.2004.00671.x
CrossRef Full Text | Google Scholar
de la Riva, E. G., Violle, C., Pérez-Ramos, I. M., Marañón, T., Navarro-Fernández, C. M. M., Olmo, M., et al. (2018). Moniulotteinen funktionaalisten ominaisuuksien lähestymistapa paljastaa ympäristöstressin jäljen Välimeren puustoisissa yhteisöissä. Ecosystems 21, 248-262. doi: 10.1007/s10021-017-0147-7
CrossRef Full Text | Google Scholar
Díaz, S., Kattge, J., Cornelissen, J. H., Wright, I. J., Lavorel, S., Dray, S., et al. (2016). Kasvien muodon ja toiminnan globaali spektri. Nature 529, 167-171. doi: 10.1038/nature16489
PubMed Abstract | CrossRef Full Text | Google Scholar
Díaz, S., Lavorel, S., de Bello, F., Quétier, F., Grigulis, K. ja Robson, T. M. (2007). Kasvien toiminnallisen monimuotoisuuden vaikutusten sisällyttäminen ekosysteemipalvelujen arviointiin. Proc. Natl. Acad. Sci. U.S.A 104, 20684-20689. doi: 10.1073/pnas.0704716104
PubMed Abstract | CrossRef Full Text | Google Scholar
Donoghue, M. J., and Edwards, E. J. (2014). Biomin siirtymät ja niche evoluutio kasveissa. Annu. Rev. Ecol. Evol. Syst. 45, 547-572. doi: 10.1146/annurev-ecolsys-120213-091905
CrossRef Full Text | Google Scholar
Douma, J. C., Aerts, R., Witte, J. P. M., Bekker, R. M., Kunzmann, D., Metselaar, K., et al. (2012). Toiminnallisesti erilaisten kasviominaisuuksien yhdistelmä tarjoaa keinon ennustaa kvantitatiivisesti monenlaisia lajikokoonpanoja Luoteis-Euroopassa. Ecography 35, 364-373. doi: 10.1111/j.1600-0587.2011.07068.x
CrossRef Full Text | Google Scholar
Engemann, K., Sandel, B., Enquist, B. J., Jørgensen, P. M., Kraft, N., Marcuse-Kubitza, A., et al. (2016). Patterns and drivers of plant functional group dominance across the Western Hemisphere: a macroecological re-assessment based on a massive botanical dataset. Bot. J. Linn. Soc. 180, 141-160. doi: 10.1111/boj.12362
CrossRef Full Text | Google Scholar
Enquist, B. J., Condit, R., Peet, R., Thiers, B., and Schildhauer, M. (2016). The botanical information and ecology network (BIEN): cyberinfrastructure for an integrated botanical information network to investigate the ecological impacts of global climate change on plant biodiversity. PeerJ Prepr. 4:e2615v2. doi: 10.7287/peerj.preprints.2615v2
CrossRef Full Text | Google Scholar
Garnier, E., Cortez, J., Billès, G., Navas, M. L., Roumet, C., Debussche, M., et al. (2004). Kasvien toiminnalliset merkkiaineet kuvaavat ekosysteemin ominaisuuksia sekundaarisen sukkession aikana. Ecology 85, 2630-2637. doi: 10.1890/03-0799
CrossRef Full Text | Google Scholar
Goldsmith, G. R., Morueta-Holme, N., Sandel, B., Fitz, E. D., Fitz, S. D., Boyle, B., et al. (2016). Plant-O-Matic: dynaaminen ja mobiili opas kaikista Amerikan kasveista. Methods Ecol. Evol. 7, 960-965. doi: 10.1111/2041-210X.12548
CrossRef Full Text | Google Scholar
Goolsby, E. W., Bruggeman, J., and Ané, C. (2017). Rphylopars: nopeat monimuuttujaiset fylogeneettiset vertailumenetelmät puuttuville tiedoille ja lajin sisäiselle vaihtelulle. Methods Ecol. Evol. 8, 22-27. doi: 10.1111/2041-210X.12612
CrossRef Full Text | Google Scholar
Grenié, M., Denelle, P., Tucker, C. M., Munoz, F., and Violle, C. (2017). funrar: R-paketti funktionaalisten harvinaisuuksien karakterisointiin. Divers. Distrib. 23, 1365-1371. doi: 10.1111/ddi.12629
CrossRef Full Text | Google Scholar
Hortal, J., de Bello, F., Diniz-Filho, J. A. F., Lewinsohn, T. M., Lobo, J. M. ja Ladle, R. J. (2015). Seitsemän puutetta, jotka vaivaavat laajamittaista tietämystä biologisesta monimuotoisuudesta. Annu. Rev. Ecol. Evol. Syst. 46, 523-549. doi: 10.1146/annurev-ecolsys-112414-054400
CrossRef Full Text | Google Scholar
Kerkhoff, A. J., Enquist, B. J., Elser, J. J. ja Fagan, W. F. (2005). Plant allometry, stoichiometry and the temperature-dependence of primary productivity. Glob. Ecol. Biogeogr. 14, 585-598. doi: 10.1111/j.1466-822X.2005.00187.x
CrossRef Full Text | Google Scholar
Kerkhoff, A. J., Fagan, W. F., Elser, J. J. ja Enquist, B. J. (2006). Fylogeneettinen ja kasvumuotovaihtelu siemenkasvien typen ja fosforin skaalautumisessa. Am. Nat. 168, E103-E122. doi: 10.1086/507879
PubMed Abstract | CrossRef Full Text | Google Scholar
Kraft, N. J., Valencia, R., ja Ackerly, D. D. (2008). Toiminnalliset piirteet ja niche-pohjainen puuyhteisön kokoaminen Amazonin metsässä. Science 322, 580-582. doi: 10.1126/science.1160662
PubMed Abstract | CrossRef Full Text | Google Scholar
Lamanna, C., Blonder, B., Violle, C., Kraft, N. J., Sandel, B., et al. (2014). Funktionaalinen ominaisuusavaruus ja leveysasteen monimuotoisuusgradientti. Proc. Natl. Acad. Sci. U.S.A. 111, 13745-13750. doi: 10.1073/pnas.1317722111
PubMed Abstract | CrossRef Full Text | Google Scholar
Lavorel, S., ja Garnier, E. (2002). Yhteisöjen koostumuksen ja ekosysteemien toiminnan muutosten ennustaminen kasvien ominaisuuksien perusteella: Pyhän Graalin maljan uudelleen tutkiminen. Funct. Ecol. 16, 545-556. doi: 10.1046/j.1365-2435.2002.00664.x
CrossRef Full Text | Google Scholar
Lavorel, S., Grigulis, K., Lamarque, P., Colace, M.-P., Garden, D., Girel, J., et al. (2011). Kasvien funktionaalisten ominaisuuksien käyttö useiden ekosysteemipalveluiden maisemallisen jakautumisen ymmärtämiseksi. J. Ecol. 99, 135-147. doi: 10.1111/j.1365-2745.2010.01753.x
CrossRef Full Text | Google Scholar
Li, Y., Shipley, B., Price, J. N., Dantas, V. de L., Tamme, R., Westoby, M., et al. (2018). Elinympäristön suodattaminen määrittää kasviyhteisöjen funktionaalisen kapean miehityksen maailmanlaajuisesti. J. Ecol. 106, 1001-1009. doi: 10.1111/1365-2745.12802
CrossRef Full Text | Google Scholar
Magallón, S., Gómez-Acevedo, S., Sánchez-Reyes, L. L., and Hernández-Hernández, T. (2015). Metakalibroitu aikapuu dokumentoi kukkivien kasvien fylogeneettisen monimuotoisuuden varhaisen nousun. New Phytol. 207, 437-453. doi: 10.1111/nph.13264
PubMed Abstract | CrossRef Full Text | Google Scholar
Maitner, B. S., Boyle, B., Casler, N., Condit, R., Donoghue, J., Durán, S. M., et al. (2018). BIEN R-paketti: työkalu botanical information and ecology network (BIEN) -tietokannan käyttämiseen. Methods Ecol. Evol. 9, 373-379. doi: 10.1111/2041-210X.12861
CrossRef Full Text | Google Scholar
Moncrieff, G. R., Bond, W. J., and Higgins, S. I. (2016). Revising the biome concept for understanding and predicting global change impacts. J. Biogeogr. 43, 863-873. doi: 10.1111/jbi.12701
CrossRef Full Text | Google Scholar
Novosolov, M., Rodda, G. H., North, A. C., Butchart, S. H. M., Tallowin, O. J. S. S., Gainsbury, A. M., et al. (2017). Population density-range size relationship revisited. Glob. Ecol. Biogeogr. 26, 1088-1097. doi: 10.1111/geb.12617
CrossRef Full Text | Google Scholar
Olson, D. M., Dinerstein, E., Wikramanayake, E. D., Burgess, N. D., Powell, G. V. N., Underwood, E. C., et al. (2001). Terrestrial Ecoregions of the world: a new map of life on earth: terrestrial ecoregions of the world: a new global map of terrestrial ecoregions provides an innovative tool for conservationing biodiversity. Bioscience 51, 933-938. doi: 10.1641/0006-3568(2001)0512.0.CO;2
CrossRef Full Text | Google Scholar
Pennington, R. T., Prado, D. E., and Pendry, C. A. (2000). Neotrooppiset kausikuivat metsät ja kvaternaariset kasvillisuuden muutokset. J. Biogeogr. 27, 261-273. doi: 10.1046/j.1365-2699.2000.00397.x
CrossRef Full Text | Google Scholar
Pennington, R. T., Richardson, J. E., and Lavin, M. (2006). Insights into the historical construction of species-rich biomes from dated plant phylogenies, neutral ecological theory and phylogenetic community structure. New Phytol. 172, 605-616. doi: 10.1111/j.1469-8137.2006.01902.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Penone, C., Davidson, A. D., Shoemaker, K. T., Di Marco, M., Rondinini, C., Brooks, T. M., et al. (2014). Puuttuvien tietojen imputointi elämänhistoriallisissa ominaisuustietoaineistoissa: mikä lähestymistapa toimii parhaiten? Methods Ecol. Toiminnallinen redundanssi ja vakaus kasviyhteisöissä. J. Veg. Sci. 24, 963-974. doi: 10.1111/jvs.12047
CrossRef Full Text | Google Scholar
Prado, D. E., and Gibbs, P. E. (1993). Lajien levinneisyysmallit Etelä-Amerikan kuivissa kausimetsissä. Ann. Mo. Bot. Gard. 80, 902-927. doi: 10.2307/2399937
CrossRef Full Text | Google Scholar
Qian, H., and Sandel, B. (2017). Alueellisten angiospermikokoonpanojen fylogeneettinen rakenne Pohjois-Amerikan leveys- ja ilmastogradienttien yli. Glob. Ecol. Biogeogr. 26, 1258-1269. doi: 10.1111/geb.12634
CrossRef Full Text | Google Scholar
Reich, P. B., Wright, I. J., Cavender-Bares, J., Craine, J. M., Oleksyn, J., Westoby, M., et al. (2003). Kasvien toiminnallisen vaihtelun evoluutio: piirteet, spektrit ja strategiat. Int. J. Plant Sci. 164, S143-S164. doi: 10.1086/374368
CrossRef Full Text | Google Scholar
Silva de Miranda, P. L., Oliveira-Filho, A., Pennington, R. T., Neves, D. M., Baker, T. R. ja Dexter, K. G. (2018). Puulaji-inventointien käyttäminen biomien kartoittamiseen ja niiden ilmastollisten päällekkäisyyksien arviointiin trooppisen Etelä-Amerikan alankoalueilla. Glob. Ecol. Biogeogr. 27, 899-912. doi: 10.1111/geb.12749
CrossRef Full Text | Google Scholar
Šímová, I., Violle, C., Kraft, N. J. B., Storch, D., Svenning, J.-C., Boyle, B., et al. (2015). Shifts in trait means and varians in North American tree assemblages: species richness patterns are loosely related to the functional space. Ecography 38, 649-658. doi: 10.1111/ecog.00867
CrossRef Full Text | Google Scholar
Šímová, I., Violle, C., Svenning, J.-C., Kattge, J., Engemann, K., Sandel, B., et al. (2018). Tärkeimpien kasviominaisuuksien alueelliset mallit ja ilmastosuhteet Uudessa maailmassa eroavat puu- ja ruohovartisten lajien välillä. J. Biogeogr. 45, 895-916. doi: 10.1111/jbi.13171
CrossRef Full Text | Google Scholar
Smith, S. A., and Beaulieu, J. M. (2009). Elämänhistoria vaikuttaa ilmastollisen kapean evoluution nopeuteen kukkivissa kasveissa. Proc. Biol. Sci. 276, 4345-4352. doi: 10.1098/rspb.2009.1176
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, S. A., and Brown, J. W. (2018). Laajasti kattavan siemenkasvien fylogenian rakentaminen. Am. J. Bot. 105, 302-314. doi: 10.1002/ajb2.1019
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, S. A., and Donoghue, M. J. (2008). Molekulaarisen evoluution nopeudet ovat yhteydessä elämänhistoriaan kukkivissa kasveissa. Science 322, 86-89. doi: 10.1126/science.1163197
PubMed Abstract | CrossRef Full Text | Google Scholar
Swenson, N. G. (2014). Phylogenetic imputation of plant functional trait databases.Ecography 37, 105-110. doi: 10.1111/j.1600-0587.2013.00528.x
CrossRef Full Text
Swenson, N. G., Enquist, B. J., Pither, J., Kerkhoff, A. J., Boyle, B., Weiser, M. D., et al. (2012). Pohjois- ja Etelä-Amerikan puuvartisten kasvien toiminnallisen monimuotoisuuden biogeografia ja suodattaminen. Glob. Ecol. Biogeogr. 21, 798-808. doi: 10.1111/j.1466-8238.2011.00727.x
CrossRef Full Text | Google Scholar
Swenson, N. G., Weiser, M. D., Mao, L., Araújo, M. B., Diniz-Filho, J. A. F., Kollmann, J., et al. (2017). Fylogenia ja puiden funktionaalisen monimuotoisuuden ennustaminen uusissa mannermaisissa ympäristöissä. Glob. Ecol. Biogeogr. 26, 553-562. doi: 10.1111/geb.12559
CrossRef Full Text | Google Scholar
Umaña, M. N., Mi, X., Cao, M., Enquist, B. J., Hao, Z., Howe, R., et al. (2017). Toiminnallisen ainutlaatuisuuden ja spatiaalisen aggregoitumisen rooli puiden harvinaisuuden selittämisessä. Glob. Ecol. Biogeogr. 26, 777-786. doi: 10.1111/geb.12583
CrossRef Full Text | Google Scholar
van Bodegom, P. M., Douma, J. C., and Verheijen, L. M. (2014). Täysin piirteisiin perustuva lähestymistapa globaalin kasvillisuuden jakautumisen mallintamiseen. Proc. Natl. Acad. Sci. U.S.A. 111, 13733-13738. doi: 10.1073/pnas.1304551110
PubMed Abstract | CrossRef Full Text | Google Scholar
Violle, C., Reich, P. B., Pacala, S. W., Enquist, B. J. ja Kattge, J. (2014). Funktionaalisen biogeografian synty ja lupaus. Proc. Natl. Acad. Sci. U.S.A. 111, 13690-13696. doi: 10.1073/pnas.1415442111
PubMed Abstract | CrossRef Full Text | Google Scholar
Violle, C., Thuiller, W., Mouquet, N., Munoz, F., Kraft, N. J. B., Cadotte, M. W., et al. (2017). Toiminnallinen harvinaisuus: poikkeavien yksilöiden ekologia. Trends Ecol. Evol. 32, 356-367. doi: 10.1016/j.tree.2017.02.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Walker, B., Kinzig, A., and Langridge, J. (1999). Kasviominaisuuksien monimuotoisuus, joustavuus ja ekosysteemien toiminta: hallitsevien ja sivulajien luonne ja merkitys. Ecosystems 2, 95-113. doi: 10.1007/s100219900062
CrossRef Full Text | Google Scholar
Westoby, M. (1998). Lehtikorkeus-siemen (LHS) kasvi-ekologinen strategiajärjestelmä. Plant Soil 199, 213-227. doi: 10.1023/A:1004327224729
CrossRef Full Text | Google Scholar
Woodward, F. I., Lomas, M. R., and Kelly, C. K. (2004). Globaali ilmasto ja kasvibiomien jakautuminen. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 359, 1465-1476. doi: 10.1098/rstb.2004.1525
PubMed Abstract | CrossRef Full Text | Google Scholar
Wright, I. J., Reich, P. B., Westoby, M., Ackerly, D. D., Baruch, Z., Bongers, F., et al. (2004). Maailmanlaajuinen lehtitalouden kirjo. Nature 428, 821-827. doi: 10.1038/nature02403
PubMed Abstract | CrossRef Full Text | Google Scholar
Xia, J., Niu, S., Ciais, P., Janssens, I. A., Chen, J., Ammann, C., et al. (2015). Kasvien fenologian ja fysiologian yhteinen kontrolli maanpäällisen bruttoprimäärätuottavuuden suhteen. Proc. Natl. Acad. Sci. U.S.A. 112, 2788-2793. doi: 10.1073/pnas.1413090112
PubMed Abstract | CrossRef Full Text | Google Scholar
Zanne, A. E., Pearse, W. D., Cornwell, W. K., McGlinn, D. J., Wright, I. J., ja Uyeda, J. C. (2018). Angiospermien funktionaalinen biogeografia: elämää ääripäissä. New Phytol. 218, 1697-1709. doi: 10.1111/nph.15114
PubMed Abstract | CrossRef Full Text | Google Scholar
Zanne, A. E., Tank, D. C., Cornwell, W. K., Eastman, J. M., Smith, S. A., FitzJohn, R. G., et al. (2014). Kolme avainta angiospermien säteilyyn pakkasympäristöihin. Nature 506, 89-92. doi: 10.1038/nature12872
PubMed Abstract | CrossRef Full Text | Google Scholar