Arviointi | Biopsykologia | Vertaileva | Kognitiivinen | Kehitys | Kieli | Yksilölliset erot | Persoonallisuus | Filosofia | Sosiaalinen |
Metodit | Tilastot | Tilastot | Kliininen | Kasvatustieteellinen | Koulutuksellinen | Teollinen | Ammatilliset kohteet | Maailmanlaajuinen psykologian |
Biologinen:Käyttäytymisgenetiikka – Evoluutiopsykologia – Neuroanatomia – Neurokemia – Neuroendokrinologia -Neurotiede – Psykoneuroimmunologia – Fysiologinen psykologia – Psykofarmakologia(Hakemisto, pääpiirteittäin)
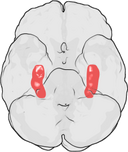
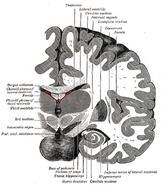
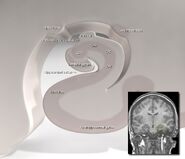
Hippokampuksen sijainti ihmisen aivoissa.
Samannimisestä aikakauslehdestä katso Hippokampus (aikakauslehti)
Hippokampus on osa etuaivoja, joka sijaitsee mediaalisessa ohimolohkossa. Se kuuluu limbiseen järjestelmään, ja sillä on merkittävä rooli pitkäkestoisessa muistissa ja avaruudellisessa navigoinnissa. Ihmisillä ja muilla nisäkkäillä on kaksi hippokampusta, yksi kummallakin puolella aivoja. Jyrsijöillä, joilla sitä on tutkittu laajimmin, hippokampus on banaanin muotoinen. Ihmisillä se on kaareva ja mutkainen, mikä muistutti varhaisia anatomeja merihevosesta. Nimi onkin peräisin kreikan kielen sanasta merihevonen (kreik. hippos = hevonen, campos = meri).
Alzheimerin taudissa hippokampus on yksi ensimmäisistä aivojen alueista, joka vaurioituu; muistihäiriöt ja sekavuus kuuluvat ensimmäisiin oireisiin. Hippokampuksen vaurioituminen voi johtua myös hapenpuutteesta (anoksiasta), enkefaliitista tai mesiaalisen ohimolohkon epilepsiasta. Henkilöillä, joilla on laaja hippokampuksen vaurio, voi esiintyä muistinmenetystä eli kyvyttömyyttä muodostaa tai säilyttää uusia muistoja.
Hippokampuksen toiminnot
Vähän varhaisin ajatus oli, että hippokampus osallistuu hajuaistimukseen: tähän näyttää viitanneen lähinnä sen sijainti aivoissa, hajuaivokuoren vieressä. Hippokampuksen hajuvasteet kiinnostavat edelleen jonkin verran, mutta lähes kukaan ei enää usko, että hippokampuksen ensisijainen tehtävä on hajuaisti.
Kirjallisuudessa on vuosien varrella vallinnut kolme pääajatusta hippokampuksen toiminnasta: estäminen, muisti ja tila. Behavioraalinen inhibitioteoria (O’Keefen ja Nadelin karrikoima “astu jarruihin!”) oli hyvin suosittu 1960-luvulle asti. Se sai suuren osan voimastaan kahdesta havainnosta: ensinnäkin eläimet, joilla on hippokampusvaurio, ovat yleensä hyperaktiivisia; toiseksi eläimillä, joilla on hippokampusvaurio, on usein vaikeuksia oppia estämään reaktioita, jotka niille on aiemmin opetettu. Jeffrey Gray kehitti tästä ajatuksesta täysimittaisen teorian hippokampuksen roolista ahdistuneisuudessa. Inhibitioteoria ei kuitenkaan ole tällä hetkellä kovin suosittu.
Toinen tärkeä ajatussuunta liittyy hippokampukseen ja muistiin. Vaikka sillä oli edeltäjiään, tämä ajatus sai päävoimansa Scovillen ja Milnerin hyvin tunnetusta raportista, joka koski hippokampuksen kirurgisen tuhoamisen tuloksia (pyrkimyksenä lievittää epileptisiä kohtauksia) potilaalla, joka tunnettiin nimellä H.M. Odottamaton lopputulos oli vakava muistinmenetys: H.M. ei kyennyt tietoisesti muistamaan tapahtumia, jotka olivat tapahtuneet hänen leikkauksensa jälkeen tai useiden vuosien ajan ennen sitä. Tapaus herätti niin valtavaa kiinnostusta, että H.M:n sanotaan nyt olevan historian intensiivisimmin tutkittu lääketieteellinen tapaus. Seuraavina vuosina on tutkittu myös muita potilaita, joilla on ollut samantasoisia hippokampusvaurioita ja muistinmenetys (onnettomuuden tai sairauden aiheuttama), ja kirjaimellisesti tuhansissa kokeissa on tutkittu hermoston plastisuuden fysiologiaa hippokampuksessa. Nykyään ollaan lähes yksimielisiä siitä, että hippokampuksella on jonkinlainen tärkeä rooli muistissa; tämän roolin täsmällisestä luonteesta keskustellaan kuitenkin edelleen laajalti.
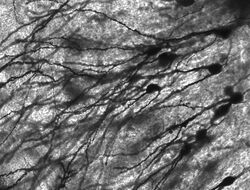
Golgi-värjäytyneet neuronit jyrsijän hippokampuksessa.
Rooli muistissa
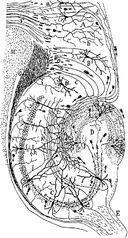
Piirros jyrsijän hippokampuksen hermopiiristä. S. Ramón y Cajal, 1911.
Psykologit ja neurotieteilijät kiistelevät hippokampuksen tarkasta roolista, mutta ovat yleensä yhtä mieltä siitä, että sillä on olennainen rooli henkilökohtaisesti koetuista tapahtumista kertovien uusien muistojen muodostamisessa (episodinen eli omaelämäkerrallinen muisti). Jotkut tutkijat pitävät hippokampusta mieluummin osana laajempaa mediaalisen ohimolohkon muistijärjestelmää, joka vastaa yleisestä deklaratiivisesta muistista (muistot, jotka voidaan eksplisiittisesti sanallistaa – näihin kuuluisi episodisen muistin lisäksi esimerkiksi faktojen muistaminen).
Pääartikkeli: Hippokampuksen rooli muistissa
Joitakin todisteita viittaa siihen, että hippokampus osallistuu avaruudellisen informaation tallentamiseen ja käsittelyyn. Rotilla tehdyt tutkimukset ovat osoittaneet, että hippokampuksen neuroneilla on spatiaalisia laukaisukenttiä. Näitä soluja kutsutaan paikkasoluiksi. Osa soluista laukeaa, kun eläin havaitsee olevansa tietyssä paikassa riippumatta kulkusuunnasta, kun taas useimmat ovat ainakin osittain herkkiä pään suunnalle ja kulkusuunnalle.
Pääartikkeli: Hippokampuksen rooli avaruudellisessa muistissa ja navigoinnissa
Hippokampus ja masennus
- Hippokampuksen tilavuushäviö,joka kenties heijastaa liiallisten glukokortikoidien vaikutuksia
neurogeneesiin, on havaittu masentuneiden ihmisten ryhmissä verrattuna kontrolliryhmiin.
Pääartikkeli: Masennus ja hippokampus
Historia
Anatomi Giulio Cesare Aranzi (noin 1564) käytti ensimmäisen kerran termiä hippokampus kuvaamaan aivojen elintä, koska se muistuttaa visuaalisesti merihevosta. Tämä elin yhdistettiin aluksi hajuaistiin eikä niinkään sen tunnettuun tehtävään muistin hankkimisessa. Venäläinen Vladimir Bekhterev havaitsi hippokampuksen roolin muistissa noin vuonna 1900 perustuen havaintoihinsa potilaasta, jolla oli syviä muistihäiriöitä. Monien vuosien ajan perinteinen käsitys hippokampuksesta oli kuitenkin se, että se oli muun limbisen järjestelmän tavoin vastuussa tunteista.
Hippokampuksen merkityksen muistissa toi tutkijoiden tietoisuuteen potilas HM. HM kärsi useista anterogradista ja ajallisesti porrastetuista retrogradista muistihäiriöistä (tällaisista häiriöistä on tehty elokuva Memento) sen jälkeen, kun häneltä oli poistettu molemminpuolisesti erilaisia mediaalisen temporaalilohkon rakenteita (mukaan lukien hippokampuksen molemminpuolinen poisto) usein esiintyvien epileptisten kohtausten lievittämiseksi. Erityisen tärkeää on, että HM kykeni edelleen oppimaan proseduraalisia tehtäviä (jotka liittyvät striatumiin) ja että hänen älykkyysosamääränsä oli keskimääräistä korkeampi. HM osoitti, että älykkyysosamäärän ja deklaratiivisen muistin välillä oli silmiinpistävä yksittäinen ero. Hippokampusmuodostuman suhteellinen koko suhteessa aivojen kokonaistilavuuteen on usein säilynyt useimmissa nisäkäslajeissa. On kuitenkin havaittu, että nämä alueet ovat suhteellisen hypotrofisia valaille.
Anatomia
Pääartikkeli: Hippokampuksen anatomia
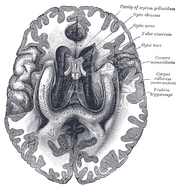
Ihmisen hippokampus.
Nissl-värjätty koronaalileikkaus makakkiapinan aivoista, jossa näkyy hippokampus (ympyröity). Lähde: brainmaps.org
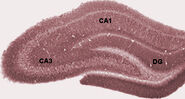
Anatomisesti hippokampus on aivokuoren reunan kehittymä. Se voidaan erottaa vyöhykkeenä, jossa aivokuori kapenee yhdeksi kerrokseksi hyvin tiheästi pakkautuneita neuroneja, joka kaartuu tiukaksi S-kirjaimen muodoksi. Aivokuoren reunaa reunustavat rakenteet muodostavat niin sanotun limbisen järjestelmän (lat. limbus = raja): niihin kuuluvat hippokampus, cingulaarinen aivokuori, hajuaisti ja amygdala. Paul MacLean ehdotti aikoinaan osana kolmiyhteistä aivoteoriaansa, että limbiset rakenteet muodostavat tunteiden hermostollisen perustan. Useimmat neurotieteilijät eivät kuitenkaan enää usko, että käsite yhtenäisestä “limbisestä järjestelmästä” on pätevä.
Hippokampus kokonaisuutena näyttää lopulta jotakuinkin kaarevalta putkelta, jota on analogisoitu eri tavoin merihevoselle, pässin sarvelle (Cornu Ammonis) tai banaanille.Tämä yleinen ulkoasu pätee kaikissa nisäkäslajeissa siilistä ihmiseen, vaikka yksityiskohdat vaihtelevat. Rotilla kaksi hippokampusta näyttää hämmästyttävän paljon banaaniparilta, jotka on liitetty yhteen varren kohdalta. Ihmisen tai apinan aivoissa hippokampuksen alhaalla oleva osa, lähellä ohimolohkon pohjaa, on paljon leveämpi kuin ylhäällä oleva osa. Yksi tämän monimutkaisen geometrian seurauksista on se, että hippokampuksen poikkileikkauksissa voi näkyä hämmentävän monenlaisia muotoja leikkauskulmasta ja -paikasta riippuen.
Hippokampuksen vahvimmat yhteydet ovat entorinaaliseen aivokuoreen (EC), joka sijaitsee sen vieressä ohimolohkossa. EK:n pinnalliset kerrokset antavat hippokampukselle eniten syötteitä, ja EK:n syvät kerrokset vastaanottavat eniten ulostuloja. EK on puolestaan vahvasti ja vastavuoroisesti yhteydessä moniin muihin aivokuoren osiin. Hippokampus saa myös erittäin tärkeän projektion mediaaliselta septaalialueelta. Septaalialueen tuhoaminen poistaa hippokampuksen theta-rytmin ja heikentää vakavasti tietyntyyppistä muistia. (Niin sanottujen “deittiraiskaushuumeiden” uskotaan aiheuttavan muistinmenetysvaikutuksensa ainakin osittain antagonisoimalla mediaalisen septumin kolinergistä projektiota hippokampukseen.)
Fysiologia
Esimerkkejä rotan hippokampuksen EEG:stä ja CA1:n neuraalisesta aktiivisuudesta theta- (hereillä/käyttäytymässä) ja LIA- (hitaasti aaltoileva uni) tiloissa. Kukin kuvaaja näyttää 20 sekunnin datan, jossa hippokampuksen EEG-jälki on ylhäällä, piikkirasterit 40 samanaikaisesti rekisteröidyistä CA1-pyramidisoluista keskellä (jokainen rasteriviiva edustaa eri solua) ja juoksunopeuden kuvaaja alhaalla. Ylin kuvaaja edustaa ajanjaksoa, jonka aikana rotta etsi aktiivisesti hajallaan olevia ruokapellettejä. Alemmassa kuvaajassa rotta nukkui.
Hippokampuksessa on kaksi pääasiallista aktiivisuuden “moodia”, joista kumpikin liittyy erilliseen EEG-aaltojen ja hermopopulaatioiden aktiivisuuden malliin.Nämä moodit on nimetty niihin liittyvien EEG-kuvioiden mukaan: theta ja suuri epäsäännöllinen aktiivisuus (LIA). Seuraavassa on joitakin niiden pääpiirteitä rotalla, eläimellä, jota on tutkittu laajimmin:
Theta-tila esiintyy aktiivisen, valppaan käyttäytymisen aikana (erityisesti liikkumisen aikana) ja myös REM-unen (unen) aikana. Theta-tilassa EEG:tä hallitsevat suuret säännölliset aallot, joiden taajuusalue on 6-9 Hz, ja hippokampuksen neuronien pääryhmät (pyramidisolut ja granulussolut) osoittavat harvaa populaatioaktiivisuutta, mikä tarkoittaa sitä, että lyhyen ajanjakson aikana suurin osa soluista on hiljaisia, kun taas jäljelle jäävä pieni osa laukeaa suhteellisen suurella nopeudella, jopa 50 piikkiä sekunnissa kaikkein aktiivisimmilla niistä. Aktiivinen solu pysyy tyypillisesti aktiivisena puolesta sekunnista muutamaan sekuntiin. Kun rotta käyttäytyy, aktiiviset solut hiljenevät ja uudet solut aktivoituvat, mutta aktiivisten solujen kokonaisprosenttiosuus pysyy jokseenkin vakiona. Monissa tilanteissa solujen aktiivisuus määräytyy suurelta osin eläimen avaruudellisen sijainnin mukaan,mutta myös muut käyttäytymismuuttujat vaikuttavat siihen selvästi.
LIA-tila esiintyy hitaiden aaltojen (ei-unien) unen aikana ja myös valveilla olevien liikkumattomuustilojen, kuten levon tai syömisen, aikana. LIA-tilassa EEG:tä hallitsevat terävät aallot, jotka ovat satunnaisesti ajoitettuja EEG-signaalin suuria poikkeamia, jotka kestävät 200-300 sekuntia. Nämä terävät aallot määrittävät myös populaation neuraaliset aktiivisuuskuviot. Niiden välissä pyramidisolut ja jyvässolut ovat hyvin hiljaisia (mutta eivät hiljaisia). Jyrkän aallon aikana jopa 5-10 % populaatiosta voi lähettää toimintapotentiaalia 50 sekunnin aikana; monet näistä soluista eivät lähetä yhtä vaan piikkien pursetta.
Nämä kaksi hippokampuksen aktiivisuusmuotoa voidaan nähdä sekä kädellisillä että rotilla, sillä tärkeällä poikkeuksella, että kädellisten hippokampuksessa on ollut vaikea nähdä voimakasta theta-rytmiikkaa. On kuitenkin olemassa laadullisesti samankaltaisia teräviä aaltoja ja samankaltaisia tilasta riippuvia muutoksia hermopopulaatioaktiivisuudessa..
Theta-rytmi
Tiheästi pakattujen hermokerrostensa vuoksi hippokampus tuottaajoitakin aivorakenteiden suurimpia EEG-signaaleja. Joissakin tilanteissa EEG:tä hallitsevat säännölliset aallot, jotka usein jatkuvat useita sekunteja. Tämä EEG-kuvio tunnetaan nimellä theta-rytmi. Se oli yksi varhaisimmista EEG-ilmiöistä, jotka löydettiin: ensimmäinen kuvaus tuli Jungilta ja Kornmullerilta vuonna 1938. Kiinnostus heräsi kuitenkin vasta vuonna 1954, kun Green ja Arduini julkaisivat pitkän ja perusteellisen tutkimuksen theta-rytmistä kaneilla, kissoilla ja apinoilla. Ehkäpä suurelta osin siksi, että he liittivät theta-rytmin kiihottumiseen, joka oli tuolloin ajankohtainen aihe, heidän artikkelinsa herätti jatkotutkimusten tulvan, jonka tuloksena julkaistiin kirjaimellisesti satoja tutkimuksia, jotka koskivat theta-rytmin fysiologiaa ja farmakologiaa 1950- ja 1960-luvuilla. Tästä melko pelottavasta työmäärästä huolimatta monet kysymykset jäivät vaille vastausta, erityisesti kysymys toiminnasta. Tähän kaikkein kriittisimpään kysymykseen ei ole vielä tälläkään hetkellä saatu vakuuttavaa vastausta.
Theta-rytmiikka on hyvin ilmeinen kaneilla ja jyrsijöillä, ja sitä esiintyy selvästi myös kissoilla ja koirilla. Se, voidaanko theta-rytmi näkyä esikoisilla, on kiusallinen kysymys. Green ja Arduini raportoivat apinoilla vain hyvin lyhyitä, melko epäsäännöllisen rytminpurkauksia, ja useimmissa myöhemmissä tutkimuksissa on havaittu vain vähän enemmän. Menetelmien vaihtelu on kuitenkin vaikeuttanut vahvojen johtopäätösten tekemistä.
Rotilla (eläimillä, joita on tutkittu ylivoimaisesti eniten) theta-taajuus näkyy pääasiassa kahdessa tilanteessa: ensinnäkin, kun eläin kävelee tai on muulla tavoin aktiivisessa vuorovaikutuksessa ympäristönsä kanssa; toiseksi, REM-unen aikana. Taajuus kasvaa juoksunopeuden funktiona, alkaen noin 6,5 Hz:stä matalalla tasolla ja nousten noin 9 Hz:iin korkealla tasolla, vaikka korkeampia taajuuksia nähdään joskus dramaattisissa liikkeissä, kuten hyppyissä laajojen aukkojen yli. Muilla, suuremmilla eläinlajeilla setataajuudet ovat yleensä hieman matalampia. Myös käyttäytymisriippuvuus näyttää vaihtelevan lajeittain: kissoilla ja kaneilla theta-taajuuksia havaitaan usein liikkumattoman valppauden aikana. Tätä on raportoitu myös rotilla, mutta vain silloin, kun ne ovat vakavasti peloissaan.
Teta ei rajoitu vain hippokampukseen. Rotilla sitä voidaan havaita monissa aivojen osissa, mukaan lukien lähes kaikissa, jotka ovat vahvassa vuorovaikutuksessa hippokampuksen kanssa. Rytmin tahdistimen uskotaan sijaitsevan mediaalisella septaalialueella: tämä alue projisoituu kaikille alueille, joilla esiintyy theta-rytmiikkaa, ja sen tuhoaminen eliminoi thetan koko aivoista. (Voi olla yksi poikkeus, pieni alue hypotalamuksessa nimeltä supramamillarynucleus, joka näyttää kykenevän ylläpitämään thetaa septumista riippumatta joissakin tilanteissa.)
Thetan funktiota, olettaen, että sillä on jokin funktio, ei ole vielä selitetty vakuuttavasti, vaikka lukuisia teorioita on ehdotettu.Suosituin suuntaus on ollut sen liittäminen oppimiseen ja muistiin.On hyvin todistettu, että mediaalisen septumin – theta-järjestelmän keskeisen solmun – vauriot aiheuttavat vakavia muistihäiriöitä.Keskimmäinen septum on kuitenkin muutakin kuin vain thetan ohjaaja, se on myös tärkein lähde kolinergisille projektioille hippokampukseen. Ei ole osoitettu, että septumvauriot aiheuttaisivat vaikutuksensa nimenomaan poistamalla theta-aaltoja.
Terävät aallot
Nukkumisen aikana tai valveillaolon aikana, kun eläin lepää tai ei muuten ole tekemisissä ympäristönsä kanssa, hippokampuksen EEG:ssä näkyy epäsäännöllisiä hitaita aaltoja, joiden amplitudi on jonkin verran suurempi kuin theta-aaltojen. Tämä kuvio keskeytyy toisinaan suurten aaltojen, niin sanottujen terävien aaltojen, takia. Näihin tapahtumiin liittyy CA3:n ja CA1:n pyramidisoluissa esiintyviä 50-100 sekunnin pituisia piikkiaktiivisuuden purskeita. Niihin liittyy myös lyhytaikaisia korkeataajuisia EEG-värähtelyjä, joita kutsutaan “aaltoiluksi”. Rippelit, joiden taajuudet ovat rotilla välillä 150-200 Hz, voidaan yleensä havaita vain elektrodeilla, jotka sijaitsevat joko CA1-solukerroksen sisällä tai hyvin lähellä sitä. Sen sijaan missä tahansa hippokampuksen sisällä tai jopa viereisissä aivorakenteissa sijaitsevat elektrodit havaitsevat usein teräviä aaltoja suurina hitaina EEG-heilahduksina, jotka kestävät 200-400 sekuntia.
Rotilla terävät aallot ovat voimakkaimpia unen aikana, jolloin niitä esiintyy keskimäärin noin yksi sekunnissa, mutta hyvin epäsäännöllisesti ajallisesti. Teräviä aaltoja esiintyy myös inaktiivisten valveillaolotilojen aikana, mutta ne ovat silloin harvinaisempia ja yleensä pienempiä. Teräviä aaltoja on havaittu myös ihmisen ohimolohkossa ja apinoiden hippokampuksessa.Apinoilla terävät aallot ovat melko voimakkaita, mutta niitä ei esiinny läheskään yhtä usein kuin rotilla.
Yksi mielenkiintoisimmista terävien aaltojen näkökohdista on se, että ne näyttävät liittyvän muistiin. Wilson ja McNaughton 1994 ja lukuisat myöhemmät tutkimukset raportoivat, että kun hippokampuksen paikkasoluilla on päällekkäisiä alueellisia ampumakenttiä (ja siksi ne usein ampuvat lähes samanaikaisina), niillä on taipumus osoittaa korreloitua aktiivisuutta käyttäytymisistuntoa seuraavan unen aikana. Tämän korrelaation voimistumisen, jota kutsutaan yleisesti reaktivaatioksi, on todettu rajoittuvan pääasiassa teräviin aaltoihin. On ehdotettu, että terävät aallot ovat itse asiassa käyttäytymisen aikana mieleen painuneiden neuraalisten aktiivisuusmallien reaktivaatioita, jotka johtuvat synaptisten yhteyksien vahvistumisesta hippokampuksen sisällä. Tämä ajatus on keskeinen osa Buzsakin ja muiden kannattamaa “kaksivaiheista muistia” koskevaa teoriaa, jonka mukaan muistot tallennetaan hippokampukseen käyttäytymisen aikana ja siirretään myöhemmin neokorteksille unen aikana: terävien aaltojen on ehdotettu ajavan Hebbin synaptisia muutoksia hippokampuksen ulostuloratojen aivokuoren kohteissa.
Evoluutio
Hippokampuksella on yleisesti ottaen samanlainen ulkomuoto kaikissa nisäkäslajeissa, alkeellisista, kuten siilistä, kaikkein “kehittyneimpiin”, kuten ihmisiin.Hippokampuksen koon ja ruumiin koon suhde kasvaa laajalti, ja se on kädellisillä kädellisillä noin kaksi kertaa niin suuri kuin siilillä. Se ei kuitenkaan kasva läheskään samaa vauhtia kuin neokorteksin ja ruumiin koon suhde.Näin ollen hippokampus vie paljon suuremman osan aivokuoren tilavuudesta jyrsijöillä kuin kädellisillä.
Hippokampuksen koon ja avaruudellisen muistin välillä on myös yleinen yhteys: kun verrataan samankaltaisten lajien välisiä lajeja keskenään, niillä lajeilla, joilla on suurempi avaruudellisen muistin kapasiteetti, on yleensä myös suuremmat tilavuudet. Tämä suhde ulottuu myös sukupuolten välisiin eroihin: lajeissa, joissa uroksilla ja naarailla on voimakkaita eroja avaruudellisen muistin kyvyssä, niillä on myös taipumus osoittaa vastaavia eroja hippokampuksen tilavuudessa
Ei-nisäkäslajeilla ei ole nisäkkäiden hippokampusta muistuttavaa aivorakennetta, mutta niillä on sitä muistuttava rakenne. Hippokampus on, kuten edellä todettiin, lähinnä aivokuoren mediaalinen reuna. Ainoastaan nisäkkäillä on täysin kehittynyt aivokuori, mutta rakenne, josta se on kehittynyt ja jota kutsutaan palliumiksi, on kaikilla selkärankaisilla, jopa kaikkein alkeellisimmilla eläimillä, kuten lampareella tai särkikalalla. Pallium jaetaan yleensä kolmeen alueeseen: mediaaliseen, lateraaliseen ja dorsaaliseen. Mediaalinen pallium muodostaa hippokampuksen esiasteen. Se ei näennäisesti muistuta hippokampusta, koska kerrokset eivät ole S-kirjaimen muotoisia tai niitä ei kiedo hammaskehä, mutta homologisuuteen viittaavat vahvat kemialliset ja toiminnalliset yhtäläisyydet. Nyt on todisteita siitä, että nämä hippokampuksen kaltaiset rakenteet osallistuvat avaruudelliseen kognitioon linnuilla, matelijoilla ja kaloilla.
Linnuilla vastaavuus on riittävän vakiintunut, joten useimmat anatomit kutsuvat mediaalista palliaalivyöhykettä “lintuhippokampukseksi”. Lukuisilla lintulajeilla on vahvat avaruudelliset taidot,erityisesti niillä, jotka kätkevät ruokaa. On näyttöä siitä, että ruokaa kätkevillä linnuilla on suurempi hippokampus kuin muilla lintulajeilla ja että hippokampuksen vaurioituminen heikentää avaruudellista muistia.
Kalojen osalta tilanne on monimutkaisempi. Teleostikaloilla (jotka muodostavat suurimman osan nykyisistä lajeista) etuaivot ovat oudosti vääristyneet muihin selkärankaisiin verrattuna. Useimmat neuroanatomit uskovat, että teleostien etuaivot ovat pohjimmiltaan kieroutuneet, kuin ylösalaisin käännetty sukka, niin että rakenteet, jotka useimmilla selkärankaisilla sijaitsevat sisäpuolella, kammioiden vieressä, ovat teleostikaloilla ulkopuolella, ja päinvastoin. Tästä seuraa muun muassa se, että tyypillisen selkärankaisen mediaalisen palliumin (“hippokampusvyöhyke”) ajatellaan vastaavan tyypillisen kalan lateraalista palliumia. Useilla kalalajeilla (erityisesti kultakaloilla) on kokeellisesti osoitettu olevan vahva avaruudellinen muistikyky, ja ne jopa muodostavat “kognitiivisia karttoja” alueista, joilla ne asuvat. On todisteita siitä, että lateraalisen palliumin vaurioituminen heikentää avaruusmuistia (pitkän matkan navigointi, kuten lohien kotiutuminen, näyttää kuitenkin perustuvan erilaisiin mekanismeihin).
Näin ollen hippokampuksen alueen rooli navigoinnissa näyttää alkaneen kaukana selkärankaisten evoluutiossa, ennen satoja miljoonia vuosia sitten tapahtunutta jakautumista. Vielä ei tiedetä, onko mediaalisella palliumilla samanlainen rooli vielä alkukantaisemmilla selkärankaisilla, kuten haikaloilla ja rauskuilla, tai jopa lampareilla ja särkikaloilla. Joillakin hyönteislajeilla ja nilviäisillä, kuten mustekaloilla, on myös voimakkaat avaruudelliset oppimis- ja navigointikyvyt, mutta ne näyttävät toimivan eri tavalla kuin nisäkkäiden avaruudellinen järjestelmä, joten toistaiseksi ei ole perusteltua syytä olettaa, että niillä olisi yhteinen evolutiivinen alkuperä; aivojen rakenteessa ei myöskään ole riittävästi samankaltaisuuksia, jotta näissä lajeissa voitaisiin tunnistaa mitään “hippokampusta” muistuttavaa.
Ks. myös
- Amygdalohippokamptomia
- Keskimmäinen etuaivokimppu
- Septal. ytimet
Viitteet & Kirjallisuusluettelo
Keskeiset tekstit
Kirjat
- Aboitiz, F, Morales D, Montiel J (2003). Nisäkkäiden isokortexin evolutiivinen alkuperä: Kohti integroitua kehityksellistä ja toiminnallista lähestymistapaa. Behav. Brain Sciences 26: 535-552.
- Amaral, D; Lavenex P (2006). “Ch 3. Hippocampal Neuroanatomy” Andersen P, Morris R, Amaral D, Bliss T, O’Keefe J The Hippocampus Book, Oxford University Press.
- Broglio, C, Gómez A, Durán E, Ocaña FM, Jiménez-Moya F, Rodríguez F, Salas C (2002). Yhteisen etuaivojen selkärankaisten suunnitelman tunnusmerkit: Specialized pallial areas for spatial, temporal and emotional memory in actinopterygian fish. Brain Res. Bull. 57: 397-399.
- Buzsáki, G (2002). Teeta-oskillaatiot hippokampuksessa. Neuron 33: 325-340.
- Buzsáki, G (2006). Rhythms of the Brain.
- Cantero, JL, Atienza M, Stickgold R, Kahana MJ, Madsen JR, Kocsis B (2003). Unesta riippuvaiset theta-oskillaatiot ihmisen hippokampuksessa ja neokorteksissa. J. Neurosci. 23: 10897-10903.
- Colombo, M, Broadbent N (2000). Onko lintujen hippokampus nisäkkäiden hippokampuksen toiminnallinen homologi?. Neurosci. Biobehav. Rev. 24: 465-484.
- Eichnbaum, H; Cohen NJ (1993). Memory, Amnesia, and the Hippocampal System, MIT Press.
- Ekstrom, AD, Kahana MJ, Caplan JB, Fields TA, Isham EA, Newman EL, Fried I (2003). Ihmisen spatiaalisen navigoinnin taustalla olevat soluverkot. Nature 425: 184-188.
- Gray, JA; McNaughton N (2000). Ahdistuksen neuropsykologia: An Enquiry into the Functions of the Septo-Hippocampal System, Oxford University Press.
- Green, JD, Arduini AA (1954). Hippokampuksen sähköinen aktiivisuus kiihottumisessa. J. Neurophysiol. 17: 533-557.
- Jacobs, LF, Gaulin SJ, Sherry DF, Hoffman GE (1990). Spatiaalisen kognition evoluutio: sukupuolispesifiset spatiaalisen käyttäytymisen mallit ennustavat hippokampuksen kokoa. PNAS 87: 6349-6352.
- Jacobs, LF (2003). Kognitiivisen kartan evoluutio. Brain Behav. Evol. 62: 128-139.
- Kirk, IJ, McNaughton N (1991). Supramammillaaristen solujen ampuminen ja hippokampuksen rytminen hidas toiminta. Neuroreport 11: 723-725.
- Maguire, EA, Gadian DG, Johnsrude IS, Good CD, Ashburner J, Frackowiak RS, Frith CD (2000). Navigointiin liittyvät rakenteelliset muutokset taksinkuljettajien hippokampuksessa. PNAS 97: 4398-4403.
- McNaughton, BL, Battaglia FP, Jensen O, Moser EI, Moser MB (2006). Polkuintegraatio ja “kognitiivisen kartan” hermostollinen perusta. Nat. Rev. Neurosci. 7: 663-678.
- Moser, EI, Kropf E, Moser M-B (2008). Paikkasolut, rasterisolut ja aivojen avaruudellinen representaatiojärjestelmä. Ann. Rev. Neurosci. 31.
- Nieuwenhuys, R (1982). An Overview of the Organization of the Brain of Actinopterygian Fishes. Am. Zool. 22: 287-310.
- O’Kane, G, Kensinger EA, Corkin S (2004). Todisteet semanttisesta oppimisesta syvässä muistinmenetyksessä: An investigation with patient H.M.. Hippocampus 14: 417-425.
- O’Keefe, J; Nadel L (1978). The Hippocampus as a Cognitive Map, Oxford University Press.
- Portavella, M, Vargas JP, Torres B, Salas C (2002). Telencephalisen pallian vaurioiden vaikutukset spatiaaliseen, temporaaliseen ja emotionaaliseen oppimiseen kultakaloilla. Brain Res. Bull. 57: 397-399.
- Rodríguez, F, Lópeza JC, Vargasa JP, Broglioa C, Gómeza Y, Salas C (2002). Spatial memory and hippocampal pallium through vertebrate evolution: insights from reptiles and teleost fish. Brain Res. Bull. 57: 499-503.
- Sainsbury, RS, Heynen A, Montoya CP (1987). Käyttäytymiskorrelaatit hippokampuksen tyypin 2 theta rotalla. Physiol. Behav. 39: 513-519.
- Scoville, WB, Milner B (1957). Viimeaikaisen muistin menetys bilateraalisten hippokampusvaurioiden jälkeen. J. Neurol. Neurosurg. Psych. 20: 11-21.
- Shettleworth, SJ (2003). Muisti ja Hippokampuksen erikoistuminen ruokaa varastoivilla linnuilla: Challenges for Research on Comparative Cognition. Brain Behav. Evol. 62: 108-116.
- Skaggs, WE, McNaughton BL, Wilson MA, Barnes CA (1996). Teta-vaiheen prekessio hippokampuksen neuronipopulaatioissa ja ajallisten sekvenssien pakkautuminen. Hippocampus 6: 149-176.
- Squire, LR (1992). Muisti ja hippokampus: synteesi rotilla, apinoilla ja ihmisillä tehdyistä havainnoista. Psych. Rev. 99: 195-231.
- Squire, LR; Schacter DL (2002). The Neuropsychology of Memory, Guilford Press.
- Vanderwolf, CH (1969). Hippokampuksen sähköinen aktiivisuus ja tahdonalainen liike rotalla. EEG & Clin. Neurophysiol. 26: 407-418.
- Vargas, JP, Bingman VP, Portavella M, López JC (2006). Telencephalon ja geometrinen tila kultakaloilla. Eur. J. Neurosci. 24: 2870-2878.
- West, MJ (1990). Hippokampuksen stereologiset tutkimukset: erilaisten lajien, mukaan lukien siilien, laboratoriojyrsijöiden, luonnonvaraisten hiirien ja ihmisten hippokampuksen osa-alueiden vertailu. Prog. Brain Res. 83: 13-36.
Papers
- Amaral DG ja Cowan WM. 1980. Hippokampusmuodostuman subkortikaaliset afferentit apinalla. Journal of Comparative Neurology. Feb 15; 189(4):573-91.
- Duvernoy, H. (2005) The Human Hippocampus, 3rd ed. Berlin: Springer-Verlag.
Lisämateriaali
Paperit
- Google Scholar
- http://www.psycheducation.org/emotion/hippocampus.htm Mahtavat aivokierrokset!!!
- BrainMaps at UCDavis hippocampus
- Artificial Hippocampus
- BrainInfo at University of Washington hier-164
- John O’Keefe & Lynn Nadel (1978) The Hippocampus as a Cognitive Map , Oxford University Press. Full Text
Lisäkuvia
Lisää kuva tähän galleriaan
ihmisen aivot: Limbinen järjestelmä
Amygdala – Cingulaarinen gyrus – Fornulaarinen gyrus – Hippokampus – Hypotalamus – Hypothalamus – Mammillaarinen runko – Nucleus accumbens – Orbitofrontaalinen aivokuori – Parahippokampuksen gyrus
|
v-d-e
Ihmisen aivot: Etuaivot (cerebrum – aivokuori – aivopuoliskot, harmaa aine) (TA A14.1.09.002-240, 301-320, GA 9.818-826) |
|||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Etelälohko |
|
||||||||||||||||
| Parietaalilohko |
|
Superolateraalinen
|
Medial/inferior
|
Kumpikin
|
|||||||||||||
| Occipitaalilohko |
|
Superolateraalinen
|
Medial/inferior
|
||||||||||||||
| Temporaalilohko |
|
Superolateraalinen
|
Medial/inferior
|
||||||||||||||
| Interlobar sulci/fissures |
|
Superolateraalinen
|
Medial/inferior
|
||||||||||||||
| Limbic lobe |
|
Muut
|
|||||||||||||||
| Insular lobe |
Insulan pitkä gyrus – Insulan lyhyt gyri – Insulan pyöreä sulcus |
||||||||||||||||
| Yleinen |
Operculum – Aivopuoliskojen navat |
||||||||||||||||
Jotkin luokittelut ovat likimääräisiä, ja jotkut Brodmannin alueet ulottuvat gyriin.
|
|||||||||||||||||
Tällä sivulla käytetään Wikipedian Creative Commons -lisensoitua sisältöä (katso tekijät).