- Introduktion
- Metoder
- Distributionsdata och bioklassificering
- Species Composition Among Biomes
- Traitdata
- Mätning av egenskapshypervolymer
- Funktionell särskiljningsförmåga och utbredning
- Funktionellt utrymme och biomlikhet
- Resultat
- Diskussion
- Kunskapsbrister
- Gemensamma mönster inom biomer
- Förhållanden mellan olika biomes
- Slutsatser
- Datatillgänglighet
- Författarbidrag
- Finansiering
- Intressekonfliktförklaring
- Acknowledgments
- Supplementary Material
Introduktion
Ekologer och biogeografer organiserar den biologiska mångfalden av landväxter i klimatmässigt bestämda biomer, med fysiognomier som kännetecknas av de dominerande arternas tillväxtformer och funktionella egenskaper (Moncrieff et al., 2016). Biomkonceptet har faktiskt en lång historia som går tillbaka till Carl Ludwig Willdenow och Alexander von Humboldt. Willdenow insåg att liknande klimat stöder liknande vegetationsformer, och Humboldt observerade det utbredda sambandet mellan växternas utbredning, fysionomi och miljöfaktorer. Sammantaget återspeglar biombegreppet antagandet att liknande miljöpåfrestningar väljer ut arter med liknande funktionella egenskaper, oberoende av deras evolutionära historia. Samtidigt är jordens existerande biom i viss utsträckning fylogenetiskt distinkta, där många/den största delen av de karakteristiska arterna är hämtade från specifika linjer som uppvisar viktiga anpassningar, inte bara till klimatselektion utan även till ytterligare påfrestningar som eld eller megaherbivorer (Woodward et al., 2004; Pennington et al., 2006; Donoghue och Edwards, 2014). Eftersom biom representerar storskaliga regelbundenheter i jordens vegetation är det av avgörande betydelse att förstå funktionella skillnader mellan biom för modellering av den globala kolcykeln och jordsystemets funktion, inklusive reaktioner på antropogena globala förändringar (Bonan m.fl, 2012; van Bodegom et al., 2014; Xia et al., 2015).
De senaste två decennierna har det skett en snabb ökning av förståelsen av den funktionella dimensionen av växternas biologiska mångfald över kontinentala skalor (Swenson et al., 2012; Lamanna et al., 2014; Šímová et al., 2018). Omfattande datasammanställningar och analysinsatser har definierat ett begränsat funktionellt egenskapsutrymme som innefattar variation i övergripande växtstorlek, bladekonomi och hydraulik, kända som funktionella egenskapsspektrum (Wright et al., 2004; Díaz et al., 2016). Funktionella egenskapsspektra definierar fysiologiska och ekologiska avvägningar som bestämmer växters livshistoriska strategier (Westoby, 1998; Reich et al., 2003; Craine, 2009) och deras inflytande på sammansättning av samhällen (Kraft et al., 2008), ekosystemets funktion (Lavorel och Garnier, 2002; Garnier et al., 2004; Kerkhoff et al., 2005; Cornwell et al., 2008) och till och med på evolutionstakten (Smith och Donoghue, 2008; Smith och Beaulieu, 2009). Den generella karaktären och användbarheten hos spektrum av funktionella egenskaper hos växter har påskyndat deras införlivande i modeller för den globala fördelningen av vegetation (van Bodegom et al., 2014), biogeokemiska kretslopp (Bonan et al., 2012) och ekosystemtjänster (Díaz et al., 2007; Cadotte et al., 2011; Lavorel et al., 2011). Få studier har dock undersökt omfattande mått på funktionell mångfald på biomskala över både dominerande och underordnade arter och livsformer.
Det finns två kontrasterande uppsättningar av förutsägelser om hur den funktionella mångfalden varierar mellan klimatmässigt och fysionomiskt skilda biom. Å ena sidan, om den biologiska mångfalden hos biomes återspeglar variationen i det tillgängliga ekologiska nischutrymmet, bör mindre taxonomiskt diversifierade biomes representera en mindre, till stor del inbäddad delmängd av det funktionella utrymme som upptas av mer diversifierade biomes. Å andra sidan, på grund av den globala karaktären hos grundläggande avvägningar i växternas struktur och funktion, och liknande selektionstryck som verkar på samma sätt över arter, kan sammansättningar som upptar olika miljöer faktiskt dela liknande områden av egenskapsutrymme (Reich et al., 2003; Wright et al., 2004; Díaz et al., 2016). Nya studier bekräftar att sambanden mellan klimat och funktionell och taxonomisk mångfald (dvs. artrikedom) är komplexa och skalberoende. I vissa fall följer den funktionella mångfalden nära klimatgradienterna, med lägre funktionell mångfald i mer varierande och extrema förhållanden (Swenson et al., 2012; de la Riva et al., 2018), och volymen av funktionellt utrymme i lokala sammansättningar expanderar med ökande taxonomisk rikedom (Lamanna et al., 2014; Li et al., 2018). Analyser av artpooler på regional skala tyder dock på att stora förändringar i artrikedom kan vara förknippade med minimala effekter på den funktionella mångfalden (Lamanna et al., 2014; Šímová et al., 2015). Dessutom kan den taxonomiska och funktionella mångfaldens svar på klimatet skilja sig åt mellan större växttillväxtformer (Šímová et al., 2018), och den relativa mångfalden av olika växtformer kan förändras avsevärt längs de klimatgradienter som definierar olika biom (Engemann et al., 2016).
Framstegen i analysen av funktionell mångfald på kontinental till global skala har delvis begränsats av brister i data för att katalogisera de taxonomiska, fördelningsmässiga, fylogenetiska och funktionella aspekterna av den biologiska mångfalden (Hortal et al., 2015). Dessutom, även med tanke på de begränsade data som finns tillgängliga, är betydande informatikutmaningar förknippade med att standardisera och integrera stora, disparata dataset som beskriver arters geografiska utbredning, funktionella egenskaper och fylogenetiska relationer (Violle et al., 2014). Här undersöker vi fördelningen av den funktionella mångfalden av växtarter över biomerna i Nord- och Sydamerika med hjälp av databasen Botanical Information and Ecology Network (BIEN; Enquist et al., 2016; Maitner et al., 2018), som sammanställer information om fördelning och funktionella egenskaper för >100 000 arter av landväxter. Specifikt undersöker vi variationen i den funktionella mångfalden och särprägeln hos biom, baserat på kartor över fördelningen av landväxtarter på en skala av 100 × 100 km rutnätsceller och ett omfattande dataset med sex funktionella egenskaper som återspeglar de viktigaste axlarna för variation i ekologiska strategier.
Vårt mål i den här studien är 3-faldigt. För det första dokumenterar vi omfattningen av de tillgängliga data som karakteriserar biomens funktionella mångfald och särprägel för att belysa bestående brister i data. För det andra, med tanke på de tillgängliga uppgifterna, karakteriserar vi fördelningen av den funktionella mångfalden inom biomes över både dominerande och underordnade tillväxtformer. Dessa analyser gör det möjligt för oss att bättre kvantifiera den funktionella särprägeln hos ett biom genom att identifiera de vanligaste funktionella strategierna hos de mest utbredda arterna inom det. För det tredje frågar vi oss om biomes faktiskt kännetecknas av funktionellt distinkta samlingar av arter, baserat på överlappningen av multidimensionella hypervolymer i funktionellt egenskapsutrymme.
Metoder
Distributionsdata och bioklassificering
För att minska effekterna av provtagningsbias som är karakteristisk för förekomstdataset som sammanställts från flera resurser använde vi BIEN 2.0-utbredningskartorna för 88 417 växtarter som är spridda i Nord- och Sydamerika (Goldsmith et al, 2016). BIEN-databasen integrerar standardiserade växtobservationer som härrör från herbariumexemplar och inventeringar av vegetationslotter. Artutbredningskartor i BIEN-databaserna uppskattades med hjälp av ett av tre tillvägagångssätt, beroende på vilka förekomster som finns tillgängliga för varje art. För arter med endast en eller två förekomster (ca 35 % av arterna) definierades det geografiska utbredningsområdet som ett kvadratiskt område på 75 000 km2 som omger varje datapunkt. De geografiska områdena för arter med tre eller fyra förekomster fastställdes med hjälp av ett konvext skrov (ca 15 % av arterna). Slutligen erhölls kartor över utbredningsområden för arter med minst fem förekomster med hjälp av Maxent-algoritmen för modellering av arters utbredning, där 19 klimatskikt användes som prediktionsvariabler och 19 rumsliga egenvektorer som filter för att begränsa modellernas överprognoser (se Goldsmith et al, 2016 för ytterligare detaljer om metodik för utbredningskartor).
Vi överlagrade BIEN 2.0-växtartskartorna på en 100 × 100 km rutnätskarta med en Lambert Azimuthal Equal Area-projektion för att erhålla en matris för närvaro/frånvaro av arter för varje rutnätscell. Baserat på Olson et al. (2001) bioklassificering, tilldelade vi varje matrisgallercell till en av biokategorierna. På grund av beräkningsbegränsningar för de efterföljande analyserna slog vi samman vissa biomes baserat på deras likheter i klimat och vegetation och litteratur för att få en bred klassificering som beskrivs i tabell 1 (exklusive Inlandsvatten, Klippor och isar samt Mangrover). Ekoregionerna Chaco och Caatinga klassificerades som Xeric Woodlands respektive Dry forest enligt Prado och Gibbs (1993), Pennington et al. (2000), Banda et al. (2016), Silva de Miranda et al. (2018).
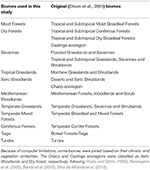
Tabell 1. Översikt över den bioklassificering som antagits i den här studien och den likvärdiga (Olson et al., 2001) bioklassificeringen (exklusive Inland Water, Rock and Ice och Mangroves).
Species Composition Among Biomes
För att förstå variationen i funktionella egenskapsutrymmen mellan olika biom utforskade vi först skillnaderna i växtartssammansättningen mellan dem. Baserat på artlistan för varje rutnätscell och biom definierade vi varje bioms karakteristiska arter som de arter som har den maximala andelen av sitt utbredningsområde i det biomet. Dessa listor över geografiskt dominerande arter i ett visst biom jämfördes med listan över arter i andra biom. Denna parvisa jämförelse ger ett enkelt sätt att härleda en riktad taxonomisk överlappning mellan biom (dvs. andelen dominerande arter i ett biom som delas med ett annat biom). Av de 88 417 arterna med tillgängliga utbredningskartor har 44 899 arter utbredningsområden som sträcker sig över mer än ett biom, och 43 518 arter är endemiska för ett specifikt biom.
Traitdata
Vi extraherade all egenskapsinformation för växtarter i Nya Världen som finns tillgänglig i datasetet BIEN 3.0 (hämtat den 7 februari 2018), vilket ger sammanlagt 80 405 observationer av egenskapsnivåer för arter. Vi filtrerade sedan informationen för sex funktionella egenskaper: maximal växthöjd (m), frömassa (mg), virkestäthet (mg/cm3), specifik bladyta SLA (cm2/g) samt bladfosfor- och bladkvävekoncentration per viktenhet (mg/g). Efter filtrering återstod totalt 18 192 observationer på artnivå. Av dessa användes en delmängd på 8 820 arter med både utbredningskartor och information om egenskaper för ytterligare analyser.
För att uppskatta det funktionella egenskapsutrymmet för varje biom krävde vi fullständiga egenskapsdata. Därför fylogenetiskt imputerade vi saknade egenskapsdata (Bruggeman et al., 2009; Penone et al., 2014; Swenson, 2014; Swenson et al., 2017) med hjälp av R-paketet “Rphylopars” v 0.2.9 (Goolsby et al., 2017) och den nyligen publicerade fylogenin för fröväxter av Smith och Brown (2018) som baslinje. Fylogenetisk imputering är ett verktyg för att förutsäga saknade data i dataset för funktionella egenskaper baserat på antagandet att närbesläktade arter tenderar att ha liknande egenskapsvärden (Swenson, 2014). Vi använde ALLBM-trädet (dvs. genbanken och Open Tree of life taxa with a backbone som tillhandahålls av Magallón et al., 2015) eftersom det maximerade överlappningen mellan arter med tillgänglig egenskaps- och distributionsinformation. Efter egenskapsimputeringen återstod totalt 7 842 arter med fullständig egenskapsinformation och utbredningskartor för vidare analys.
Mätning av egenskapshypervolymer
Vi mätte den relativa funktionella mångfalden av biom genom att beräkna egenskapshypervolymer från artpooler inom biomceller. På grund av beräkningsbegränsningar konstruerade vi egenskapshypervolymerna med hjälp av ett slumpmässigt urval av 20 % av cellerna i varje biom. Hypervolymerna för varje rutnätscell uppskattades med hjälp av de extraherade sex funktionella dragen och R-paketet “hypervolume” (Blonder et al., 2014, 2018), med hjälp av den Gaussiska KDE-metoden med standard Silverman-bandbreddsskattaren. Frömassa, höjd och virkestäthet var logtransformerade, medan SLA var kvadratrotstransformerad. Alla egenskaper skalades och centrerades före analysen. Hypervolymer rapporteras i enheter av standardavvikelser (sd) till potensen till antalet använda egenskaper (dvs. sd6).
Funktionell särskiljningsförmåga och utbredning
Eftersom biom karakteriseras av sin dominerande vegetation undersökte vi också den geografiska allmänheten och den funktionella särskiljningsförmågan hos arter inom biom. Med hjälp av kartor över arters utbredningsområde och information om funktionella egenskaper uppskattade vi varje arts funktionella särart och utbredning i varje biom, enligt det konceptuella ramverket för funktionell sällsynthet av Violle et al. (2017). Med hjälp av hela uppsättningen (uppmätta och imputerade) egenskaper mätte vi först det euklidiska avståndet i det standardiserade egenskapsutrymmet mellan alla artpar. Vi beräknade sedan den funktionella distinktiviteten (Di) för varje art i varje biom som det genomsnittliga funktionella avståndet mellan en art och de N andra arterna inom biomens artpool. Di skalades mellan 0 och 1 inom varje biom där lägre värden representerar arter som är funktionellt gemensamma (redundanta) och högre värden representerar arter som är funktionellt särskiljande, jämfört med de andra arterna i biomen. Vi uppskattar också den geografiska utbredningen (Wi) av en art i ett biom, mätt som antalet rutnätsceller som upptas av arten inom ett biom i förhållande till det totala antalet celler i det biomet. Ett värde på 1 anger att arten i fokus finns i alla rutnätsceller som täcks av biomet. Både funktionell distinktivitet och geografisk utbredning beräknades med hjälp av R-paketet “funrar” (Grenié et al., 2017).
Då vi saknar heltäckande mått på dominans baserade på lokal abundans eller biomassa använde vi måtten Di och Wi för att för varje biom identifiera de “vanligaste” arterna som de arter som både är geografiskt utbredda (Wi > 0,5) och funktionellt likartade (Di < 0,25). Det sistnämnda tröskelvärdet användes eftersom den tredje kvantilen av värdena för funktionell distinktivitet bland biomerna varierade mellan 0,2 och 0,3. För att skilja funktionellt distinkta arter från funktionellt likartade arter i efterföljande analyser använde vi därför ett värde på 0,25 som gränsvärde (dvs, arter med distinktivitetsvärden < 0,25 ansågs vara funktionellt gemensamma eller redundanta inom ett specifikt biom).
Funktionellt utrymme och biomlikhet
För att uppskatta överlappningen mellan biomers hypervolymer använde vi Sørensens likhetsindex med hjälp av (i) det totala antalet arter och (ii) listan över arter som ansågs vara funktionellt gemensamma och geografiskt utbredda för varje biom. Likheten mellan funktionella egenskaper beräknades som den parvisa fraktionella överlappningen av hypervolymerna mellan olika biom. Den fraktionella överlappningen beräknades genom att dela den dubbla volymen av skärningspunkten mellan två hypervolymer med volymen av deras föreningar. Alla hypervolymer uppskattades med hjälp av R-paketet “hypervolume” (Blonder et al., 2014, 2018), med hjälp av den Gaussiska KDE-metoden med Silvermans standardbandbreddskalkylator.
Resultat
Vi fann en betydande överlappning i sammansättningen av växtarter mellan olika biom, baserat på skärningspunkten mellan de modellerade arternas utbredningsområden (se figur 1 och kompletterande tabell 1). Denna taxonomiska överlappning är större inom tropiska och tempererade biom, med relativt få arter som delas mellan dessa två klimatzoner. Intressant nog delar xeriska skogsmarker arter med både tropiska och tempererade biom. Trots den höga andelen arter som kännetecknar tropisk fuktig skog (83 %) delar denna biom också ett stort antal arter med andra tropiska biomes, t.ex. torra skogar, savanner, xeriska skogar och tropiska gräsmarker. Den stora taxonomiska överlappningen av tempererade biomes som tempererade gräsmarker, medelhavsskogar, taiga och tundra tyder på en potentiell låg funktionell distinktivitet, där vissa tempererade biomes representerar en sämre delmängd av de mer artrika, funktionellt diversifierade tropiska biomes.
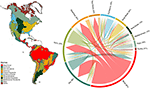
Figur 1. Överlappning av växtarter mellan biom i Nya världen. Procentvärdena uttrycker andelen arter som förekommer i ett biom och som har den största delen av sitt geografiska utbredningsområde i det biomet, N = 84 413. Basen på varje grenad pil är placerad så att den visar det biom som omfattar den största andelen av en arts utbredningsområde, medan bredden representerar det antal arter som delas med de biom som finns i pilens spetsar. Se kompletterande tabell 1 för den underliggande matrisen.
Med mindre än 10 % av de kartlagda arterna representerade var egenskapsdata ganska sparsamma för våra biometriska artsammansättningar (figur 2A). Dessutom varierade de tillgängliga egenskapsdata avsevärt mellan olika egenskaper, växtklasser och biom. Fylogenetiskt sett var blad-P och vedtäthet särskilt dåligt representerade och hela klasser saknade tillgängliga data. Som ett resultat av att av de cirka 2/3 av egenskapsvärdena för våra 7 842 arter som måste imputeras måste många tilldelas i avsaknad av närbesläktade taxa. Geografiskt sett är SLA, höjd och frömassa undermåliga i tropikerna och det tempererade Sydamerika, medan blad-N och vedtäthet är mer jämnt fördelade mellan regionerna. Blad-P är dåligt samplat i alla biomes. Hypervolymer av egenskaper som skapats från artpooler med ett slumpmässigt urval av 20 % av cellerna för varje biom visar huvudsakligen skillnader mellan tropiska och tempererade/kalla biom (figur 2B). Hypervolymer av egenskaper tenderar att vara större och mer varierande i tropiska biom, med den största variationen i fuktiga och torra tropiska skogar. Bland de tempererade/kalla biomena har barrskog och tempererad blandskog de största hypervolymerna av egenskaper. Intressant nog uppvisar xeriska buskmarker en fördelning av hypervolymer som mer liknar tropiska än tempererade biom.
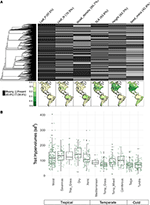
Figur 2. (A) Andel arter i BIEN 3.0-databasen med kända (grå) eller saknade (svarta) egenskapsvärden. Fylogenin till vänster motsvarar fröväxternas ALLBM-träd från Smith och Brown (2018) (dvs. GenBank och Open Tree of life taxa med en ryggrad som tillhandahålls av Magallón et al., 2015). Kartorna representerar andelen arter med egenskapsinformation i förhållande till det totala antalet arter i BIEN 2.0-databasen. (B) Fördelning av hypervolymer av egenskaper för 20 % av slumpmässigt utvalda 100 × 100 km-celler i varje biom. Hypervolymerna rapporteras i enheter av standardavvikelser till potensen av antalet använda egenskaper.
Sambandet mellan funktionell distinktivitet och geografisk spridning är anmärkningsvärt likartat mellan olika biom (figur 3). I varje biom är den stora majoriteten av arterna både geografiskt begränsade och funktionellt likartade. Samtidigt var de mest funktionellt särskiljande arterna inom varje biom i allmänhet också geografiskt begränsade, och de mest geografiskt utbredda arterna var nästan alltid funktionellt likartade.
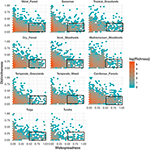
Figur 3. Mönster av funktionell distinktivitet i olika biom. Distinktivitet representerar hur arter är funktionellt avlägsna från varandra inom ett biom (dvs. det genomsnittliga parvisa fenotypiska avståndet från en fokalart till alla andra). Ju större värdet är, desto mer avlägsen är en art från centrum av biomens funktionella utrymme. Utbredning mäter hur geografiskt vanlig en art är inom ett biom. Ett värde på 0 anger att arten förekommer i en enda biocell. Bin-värden är antalet arter på en loge-skala. Rektanglar representerar de arter som anses vara funktionellt likartade och utbredda i varje biom med hjälp av gränsvärden på 0,25 och 0,5 för distinktivitet respektive utbreddhet.
Vi fann en avsevärd variation i den proportionella fördelningen av tillväxtformer inom och mellan biom. Vedartade arter utgör de dominerande tillväxtformerna i tropiska biom i termer av artantal, medan örtartade arter dominerar i tempererade miljöer (figur 4). När vi endast tog hänsyn till de arter som är utbredda och funktionellt vanliga i varje biom förändrades fördelningen av tillväxtformerna mellan olika biom, särskilt när det gäller andelen träd, örter och gräs. I tropiska biom minskade till exempel andelen träd i varje biom utom i fuktig skog. I tempererade biomes ökade andelen gräs, särskilt i polarområdena. Intressant nog liknar fördelningen av växtformer i xeriska skogar mer fördelningen i tempererade biom när endast utbredda och funktionellt vanliga arter beaktas.
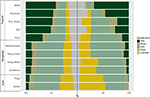
Figur 4. Fördelning av tillväxtformer i varje biom (till vänster) med hjälp av det totala antalet arter och (till höger) med hjälp av endast de arter som är funktionellt likartade och utbredda inom biomet (se figur 3).
Parvisa jämförelser av artsammansättningen mellan olika biom avslöjar tre huvudkluster som representerar de tropiska, tempererade och polära klimatzonerna (figur 5A), vilket avspeglar den höga taxonomiska överlappningen inom, men inte mellan, dessa regioner (figur 1). Parvisa jämförelser av hypervolymer av egenskaper mellan olika biom visar en mindre tydlig klustring av klimatzoner (figur 5B). I detta fall överlappar det funktionella utrymmet för xeriska skogsmarker avsevärt med tempererade biom. Denna analys visar också att taigas funktionella utrymme överlappar med tempererade gräsmarker och blandskogar. Den parvisa jämförelsen av hypervolymer av egenskaper mellan olika biom med hjälp av endast de arter som anses vara funktionellt vanliga och utbredda visar mindre överlappning av egenskapsutrymmen mellan och inom klimatzoner (figur 5C). Även om xeriska skogsmarker nu är grupperade med resten av tropiska biom, uppvisar dessa livsmiljöer tillsammans med tropiska gräsmarker stor överlappning i funktionellt utrymme med tempererade biom som medelhavsskogar och tempererade gräsmarker.
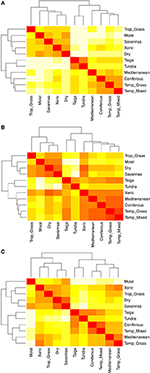
Figur 5. (A) Parvis olikhet i artsammansättning mellan olika biom. (B) Parvis olikhet i hypervolymer av egenskaper (1-Sørensen-likhet) mellan biom med hjälp av det totala antalet arter. (C) Parvis olikhet i hypervolymer av egenskaper (1-Sørensens likhet) mellan olika biom med hjälp av endast de arter som anses vara funktionellt likartade och utbredda. Ju ljusare cell desto större olikhet.
Diskussion
Våra analyser ger tre viktiga insikter för att förstå terrestra biomes genom en funktionell lins. För det första visar vi att trots framsteg i sammanställningen och syntesen av primära uppgifter om biologisk mångfald kvarstår betydande kunskapsbrister som kan begränsa vår förmåga att kvantifiera den funktionella biologiska mångfalden hos biomes på kontinental till global skala. För det andra visar våra analyser av de tillgängliga uppgifterna ändå att alla biom i Nord- och Sydamerika har ett anmärkningsvärt gemensamt mönster där de geografiskt mest utbredda arterna i ett biom tenderar att dela gemensamma funktionella egenskaper medan de mest funktionellt särskiljande arterna alltid är begränsade i sin utbredning. För det tredje, när endast de utbredda och funktionellt gemensamma arterna beaktas, kan biomes lättare särskiljas funktionellt, och mönstren av olikheter mellan biomes tycks återspegla en korrespondens mellan klimatet och växternas funktionella nischutrymme. Sammantaget tyder våra resultat på att även om studiet av den funktionella mångfalden av biom fortfarande befinner sig i ett uppbyggnadsskede, kommer en fortsatt utveckling av fältet sannolikt att ge insikter som kopplar samman evolution, biogeografi, sammansättning av samhällen och ekosystemens funktion.
Kunskapsbrister
BIEN-databasen är särskilt utformad för att täppa till luckor i vår kunskap om växters biologiska mångfald, men som Hortal et al. (2015) påpekar, leder samverkande kunskapsbrister till osäkerhet när det gäller att kvantifiera biologisk mångfald i de största skalorna. Till exempel hade inte bara färre än 10 % av våra kartlagda arter både egenskapsdata och fylogenetisk information tillgänglig, utan de saknade egenskapsdata (~66 % av arternas egenskapsvärden) var ganska strukturerade, både fylogenetiskt och geografiskt. Den Raunkiaeriska bristen på funktionella egenskapsdata (sensu Hortal et al., 2015) samverkar alltså med den Wallaceanska bristen på fördelningsinformation och den Darwinska bristen på fylogenetisk förståelse. Och även om vi kan använda fylogenetisk kunskap för att på ett rimligt sätt ersätta dessa saknade värden (Swenson et al., 2017), förblir flera aspekter av de resulterande mönstren som är viktiga för bioklassificering osäkra. Till exempel tyder de befintliga uppgifterna om tillväxtformer på att vedartade arter dominerar i alla tropiska biom, medan den proportionella mångfalden av örtartade tillväxtformer är mycket högre i tempererade och polära biom (se även Engemann et al., 2016; Zanne et al., 2018). Avspeglar denna latitudinella ökning av den relativa mångfalden av örtartade växter provtagningsbias och bristande taxonomisk kunskap om tropiska örtartade växter, eller återspeglar den de olika evolutionära och biogeografiska historierna i de Nearktiska och Neotropiska rikena? Dominerande växtformer är en av de viktigaste särskiljande egenskaperna hos biom, så systematiska förändringar i de mest diversifierade växtformerna (oavsett om de är dominerande eller inte) kommer med nödvändighet att påverka våra förutsägelser om hur den funktionella mångfalden kan förändras i olika biom, och hur dessa förändringar kommer att påverka ekosystemens funktion och tjänster.
Vi använde kartor över arters utbredningsområden som indata, främst för att övervinna det obestridliga problemet med provtagningsbias som är karakteristiskt för datamängder som sammanställts från flera olika källor (se tilläggsfigur 1). Utbredningskartor som är hämtade från modeller för arters utbredning kan dock utgöra ytterligare ett element av osäkerhet i våra resultat. Med tanke på den geografiska upplösningen i den här studien och den rumsliga komplexiteten hos vissa biom kan dessa modeller till exempel ha överskattat vissa arters geografiska utbredning från celler i ett enda biom till celler i närliggande och sammanhängande biom. Användningen av uppgifter om förekomst förändrade inte de allmänna resultaten av denna studie (se de kompletterande figurerna 2-6), men en liten minskning av den funktionella överlappningen mellan biomerna skulle faktiskt kunna avslöja en överskattning av vissa arters utbredning (se de kompletterande figurerna 2, 5 och 6). Användningen av information om förekomst förändrade också de allmänna förhållandena mellan funktionell distinktivitet och utbredning i våra resultat, på grund av den relativa överprovtagningen av tempererade jämfört med tropiska regioner (se kompletterande figur 4). I detta fall begränsas vårt mått på utbredning av den extremt begränsade provtagningen av de flesta arter i tropiska biom. I takt med att dataset och provtagning förbättras kommer det sannolikt att finnas utrymme för att minska dessa osäkerheter i framtiden.
Den andra datafrågan som måste åtgärdas för att förstå den funktionella mångfalden i biom är den Prestoniska bristen på information om abundans. Abundans är särskilt viktigt i studier av funktionell mångfald eftersom de egenskaper som har störst betydelse för samhällets sammansättning, ekosystemets funktion och biogeokemiska kretslopp är egenskaperna hos de mest dominerande arterna. Eftersom vi baserade våra sammansättningar på kartor över utbredningsområden som härrörde från modeller för artutbredning kunde vi bara ta upp den geografiska komponenten av allmän spridning, men det positiva förhållandet mellan lokal abundans och geografisk utbredning, det s.k. ockupations-abundansförhållandet, tyder på att de mest utbredda arterna i varje biom ofta också kan vara bland de mer rikliga (Borregaard och Rahbek, 2010; Novosolov et al., 2017). Eftersom biomes i allmänhet kännetecknas av de dominerande växtformerna i regionen kan integrering av information om abundans resultera i biomes som uppvisar mindre funktionell överlappning. Även om BIEN har sammanställt omfattande data om parceller som gör det möjligt att uppskatta den lokala abundansen, försvåras dock rimliga jämförelser mellan olika regioner och tillväxtformer av ojämn provtagning, regionala skillnader i gammadiversitet och inkommensurabla metoder för att kvantifiera abundansen.
Gemensamma mönster inom biomer
Trots de ihållande luckorna i de tillgängliga uppgifterna avslöjade våra analyser ett tidigare odokumenterat förhållande som är gemensamt för alla biomer på västra halvklotet: i alla biomer tenderar de mest utbredda arterna också att uppvisa låg funktionell särskiljningsförmåga, medan de mest funktionellt särskiljande arterna nästan undantagslöst är begränsade i sin utbredning (figur 3). Detta förhållande mellan “beläggning och redundans” kan tyda på att de klimatförhållanden som råder inom ett biom väljer ut en uppsättning gemensamma egenskaper, där mer funktionellt distinkta arter är begränsade till en mer sällsynt delmängd av livsmiljöer inom biomet. Den övergripande förekomsten av funktionellt likartade arter i alla biom och “ockupation-redundans”-förhållandet stämmer båda överens med en nyligen genomförd global analys av funktionell mångfald på samhällsnivå som tyder på att filtrering av livsmiljöer leder till samexistens av funktionellt likartade arter (Li et al., 2018) samt med studier som visar att funktionella redundanser ökar samhällets stabilitet (t.ex. Walker et al., 1999; Pillar et al., 2013). Tillsammans med den höga graden av både taxonomisk och funktionell överlappning mellan olika biom (figurerna 1 och 5) förstärker dessutom det faktum att vanliga, utbredda arter är funktionellt likartade uppfattningen att landväxter över ett brett spektrum av miljöförhållanden har gemensamma egenskaper nära kärnan av det funktionella egenskapsspektrumet (Wright et al., 2004; Díaz et al., 2016). Till skillnad från Umaña et al. (2017) visar våra resultat att sällsynta, geografiskt begränsade arter kan eller inte kan vara funktionellt skilda från mer utbredda och vanliga arter. Frågan är alltså om mer funktionellt distinkta arter är specialister i vissa miljöer eller om deras kombinationer av egenskaper resulterar i demografiska eller fysiologiska kompromisser som begränsar deras geografiska utbredning.
Förhållanden mellan olika biomes
Vår jämförelse av fördelningen av hypervolymen av egenskaper mellan olika biomes (figur 2B) är förenlig med observationen att mer artrika miljöer också är mer funktionellt diversifierade (Swenson m.fl, 2012; Lamanna et al., 2014; Li et al., 2018; Šímová et al., 2018). Våra resultat är mer tvetydiga när det gäller hypotesen att säsongsbundna och extrema klimatmiljöer konsekvent begränsar arters funktionella mångfald (de la Riva et al., 2018). Alla tropiska biom uppvisar högre genomsnittlig funktionell mångfald än alla tempererade biom, och de polära biom uppvisar faktiskt de minsta hypervolymerna. Inom varje grupp uppvisar dock torrare eller mer säsongsvariabla biomes inte alltid mindre hypervolymer (t.ex. torra skogar, xeriska skogsmarker), vilket vi skulle ha förväntat oss enligt toleranshypotesen av Currie et al. (2004), medan optimala klimatförhållanden stöder fler kombinationer av fysiologiska parametrar. Utifrån enbart dessa resultat kan vi inte avgöra om tempererade och polära biomes är mindre taxonomiskt diversifierade på grund av begränsningar av funktionellt nischutrymme, eller om deras funktionella hypervolymer är små på grund av att de inte är taxonomiskt diversifierade. Nollmodelleringsmetoder skulle eventuellt kunna hjälpa till att särskilja taxonomisk och funktionell mångfald (Swenson et al., 2012; Lamanna et al., 2014; Šímová et al., 2018), men en sådan analys låg utanför ramen för den här studien. Viktigare är att våra resultat förstärker vikten av att förstå hur evolutionär och biogeografisk historia formar biomens funktionella mångfald (Woodward et al., 2004; Pennington et al., 2006; Donoghue och Edwards, 2014; Moncrieff et al., 2016). Det omfattande delandet av arter (och högre släktskap) mellan olika biom inom, men i stort sett inte mellan, olika biogeografiska områden skulle kunna tjäna både till att homogenisera den funktionella mångfalden inom områden och till att ge ledtrådar om de karakteristiska egenskaper som väljs för, eller mot, olika miljöer (Douma et al, 2012; Zanne et al., 2014, 2018).
Trots betydande hypervolymöverlappning mellan alla biom (kompletterande figur 2) verkar tropiska, tempererade och kalla biom alla uppta distinkta regioner av det funktionella utrymmet (figurerna 5B,C). De viktigaste egenskaperna som skiljer biomerna åt verkar vara egenskaper som är relaterade till den totala växtstorleken, inklusive både mogen höjd och frömassa (kompletterande figurer 7-10), snarare än egenskaper som rör bladekonomi, vilket observerats i mer lokala, plotbaserade analyser (t.ex. Douma et al., 2012). Undantaget är blad-P, som uppvisade betydande skillnader mellan tropiska och tempererade/polära biom (kompletterande figurer 7-10), ett mönster som har observerats i andra analyser baserade på både artspecifika värden och mätningar av hela ekosystem (Kerkhoff et al., 2005; Swenson et al., 2012). Eftersom blad-P var vår mest sparsamt provtagna egenskap kan skillnaderna mellan tropiska och tempererade/polära områden vara föremål för bias från imputeringsförfarandet. Leaf P uppvisar dock en betydande fylogenetisk signal (Kerkhoff et al., 2006), vilket tyder på att imputeringarna bör vara opåverkade (Swenson et al., 2017). Leaf P är också signifikant högre i örtartade växtformer (Kerkhoff et al., 2006), så den latitudinella förskjutningen från övervägande vedartad till örtartad mångfald (figur 4) kan också påverka detta mönster.
När vi begränsade hypervolymanalyserna till endast de mest utbredda och funktionellt vanligaste arterna i varje biom, överlappade de enskilda biomen inom varje biogeografiskt område mindre i funktionella egenskapsutrymmen (jämför figurerna 5B,C), vilket tyder på att dessa arter kan återspegla fenotyper som är bättre anpassade till särskilda miljöer. I detta sammanhang är den xeriska skogsbiomen särskilt intressant. Delvis på grund av att ekoregionen Chaco ingår, grupperar sig xeriska skogsmarker taxonomiskt med de tropiska biomen (figur 5A). Men när de funktionella hypervolymerna för alla arter beaktas visar de mycket större likhet med de tempererade biomena (figur 5B). Slutligen, när endast de mest utbredda och funktionellt vanligaste arterna analyseras, uppvisar xeriska skogsmarker återigen ökad likhet med tropiska biom, men de bibehåller också en hög likhet med tempererade gräsmarker och Medelhavsbiom (figur 5C). Övergångar från varma, mesiska miljöer till kallare, torrare och mer säsongsbetonade miljöer underlättas av liknande egenskaper, t.ex. örtrikedom (Douma et al., 2012; Zanne et al., 2014, 2018), och i likhet med kallare miljöer tenderar samhällen i torra miljöer också att vara mer fylogenetiskt klustrade (Qian och Sandel, 2017). Dessutom gör xeriska zoners läge vid gränsen för den intertropiska konvergenszonen dem till en geografisk övergång mellan de tropiska och tempererade områdena. Det faktum att xeriska skogsmarker ligger mellan de tropiska och tempererade områdena både funktionellt och biogeografiskt stärker ytterligare idén att vi för att bättre förstå den funktionella mångfalden av biom måste ta hänsyn till deras biogeografiska och fylogenetiska historia (Pennington et al., 2006; Donoghue och Edwards, 2014; Moncrieff et al, 2016).
Slutsatser
Alla klassificeringar av terrestriska biom påtvingar ett litet antal diskreta kategorier på kontinuerliga gradienter i klimat och artutbredning, och representerar därmed en grov förenkling av det komplexa ekologiska landskapet (Moncrieff et al., 2016, men se Silva de Miranda et al., 2018). Men trots deras potential för överdriven förenkling är biomes användbara konstruktioner för att organisera och förstå den biologiska mångfalden och funktionen hos jordens viktigaste terrestra ekosystem, och egenskapsbaserade tillvägagångssätt har stor potential att hjälpa till att dynamiskt modellera globala vegetationsfördelningar.
I den här studien har vi visat att en stor del av den taxonomiska mångfalden i alla biomes representerar arter som är både snävt fördelade och funktionellt likartade. Vidare tenderar de mest funktionellt särskiljande arterna inom varje biom också att vara geografiskt sällsynta, medan de utbredda arterna på ett enhetligt sätt uppvisar en låg funktionell särskiljningsförmåga. Trots en omfattande taxonomisk och funktionell överlappning mellan biomerna, grupperar de sig i distinkta, biogeografiskt och klimatmässigt skilda enheter, särskilt när den funktionella grupperingen baseras på de mest utbredda och funktionellt vanligaste arterna i varje biom. För att främja en funktionell förståelse av biom kommer det dock att krävas inte bara en bättre karakterisering av egenskapsvariation inom och mellan vegetationstyper, utan även information om artgruppernas biogeografiska och fylogenetiska historia och arternas relativa abundans inom biom.
Datatillgänglighet
Datasetterna för egenskaper och kartorna över arters utbredningsområde som analyserats för den här studien kan laddas ner via R-paketet BIEN (Maitner et al., 2018). Se http://bien.nceas.ucsb.edu/bien/ för mer information om BIEN-databasen.
Författarbidrag
SE-L, AK, BJE, DMN och CV utformade studien; SE-L och AK analyserade data; SE-L och AK skrev den första versionen av manuskriptet; CV, BB, NK, BSM, BM, RKP, BS, SS, J-CS och SKW bidrog till sammanställning och skapande av egenskaps- och utbredningskartdatabasen. Alla författare bidrog till utvecklingen och skrivandet av manuskriptet.
Finansiering
SE-L, DMN och AK stöddes av ett forskningsanslag för samarbete från US National Science Foundation (DEB-1556651). BJE fick stöd av National Science Foundation award DEB-1457812 och Macrosystems-1065861. CV fick stöd från Europeiska forskningsrådet (Grant ERC-StG-2014-639706-CONSTRAINTS) och från den franska stiftelsen för forskning om biologisk mångfald (FRB; www.fondationbiodiversite.fr) inom ramen för CESAB-projektet Causes and consequences of functional rarity from local to global scales. J-CS anser att detta arbete är ett bidrag till hans VILLUM Investigator-projekt Biodiversity Dynamics in a Changing World som finansieras av VILLUM FONDEN (bidrag 16549). SKW fick stöd från Strategic Science Investment Fund från Nya Zeelands ministerium för näringsliv, innovation och sysselsättning, Science and Innovation Group.
Intressekonfliktförklaring
Författarna förklarar att forskningen utfördes i avsaknad av kommersiella eller finansiella relationer som skulle kunna tolkas som en potentiell intressekonflikt.
Acknowledgments
Vi tackar Dan Griffith och två granskare för deras konstruktiva kritik som förbättrade det här manuskriptet avsevärt. Vi tackar också alla som bidragit med BIEN-data (se http://bien.nceas.ucsb.edu/bien/data-contributors/ för en fullständig lista).
Supplementary Material
Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2018.00219/full#supplementary-material
Banda, R.K., Delgado-Salinas, A., Dexter, K. G., Linares-Palomino, R., Oliveira-Filho, A., Prado, D., et al. (2016). Växtdiversitetsmönster i neotropiska torrskogar och deras konsekvenser för bevarandet. Science 353, 1383-1387. doi: 10.1126/science.aaf5080
PubMed Abstract | CrossRef Full Text | Google Scholar
Blonder, B., Lamanna, C., Violle, C. och Enquist, B. J. (2014). Den n-dimensionella hypervolymen. Glob. Ecol. Biogeogr. 23, 595-609. doi: 10.1111/geb.12146
CrossRef Full Text | Google Scholar
Blonder, B., Morrow, C. B., Maitner, B., Harris, D. J., Lamanna, C., Violle, C., et al. (2018). Nya metoder för att avgränsa n-dimensionella hypervolymer. Methods Ecol. Evol. 9, 305-319. doi: 10.1111/2041-210X.12865
CrossRef Full Text | Google Scholar
Bonan, G. B., Oleson, K. W., Fisher, R. A., Lasslop, G. och Reichstein, M. (2012). Att förena bladfysiologiska egenskaper och data om takflöden: användning av databaserna TRY och FLUXNET i gemenskapens markmodell version 4. J. Geophys. Res. Biogeosci. 117:G02026. doi: 10.1029/2011JG001913
CrossRef Full Text | Google Scholar
Borregaard, M. K., and Rahbek, C. (2010). Kausalitet i förhållandet mellan geografisk spridning och artrikedom. Q. Rev. Biol. 85, 3-25. doi: 10.1086/650265
PubMed Abstract | CrossRef Full Text | Google Scholar
Bruggeman, J., Heringa, J. och Brandt, B. W. (2009). PhyloPars: uppskattning av saknade parametervärden med hjälp av fylogeni. Nucleic Acids Res. 37, W179-W184. doi: 10.1093/nar/gkp370
PubMed Abstract | CrossRef Full Text | Google Scholar
Cadotte, M. W., Carscadden, K. och Mirotchnick, N. (2011). Bortom arter: funktionell mångfald och upprätthållandet av ekologiska processer och tjänster. J. Appl. Ecol. 48, 1079-1087. doi: 10.1111/j.1365-2664.2011.02048.x
CrossRef Full Text | Google Scholar
Cornwell, W. K., Cornelissen, J. H., Amatangelo, K., Dorrepaal, E., Eviner, V. T., Godoy, O., et al. (2008). Växtartsegenskaper är den dominerande kontrollen av nedbrytningshastigheten för avfall inom biom i hela världen. Ecol. Lett. 11, 1065-1071. doi: 10.1111/j.1461-0248.2008.01219.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Craine, J. M. (2009). Resursstrategier hos vilda växter. New Jersey, NJ: Princeton University Press. doi: 10.1515/9781400830640
CrossRef Full Text | Google Scholar
Currie, D. J., Mittelbach, G. G., Cornell, H. V., Field, R., Guégan, J. F., Hawkins, B. A., et al. (2004). Prediction and tests of climate-based hypotheses of broad-scale variation in taxonomic richness. Ecol. Lett. 12, 1121-1134. doi: 10.1111/j.1461-0248.2004.00671.x
CrossRef Full Text | Google Scholar
de la Riva, E. G., Violle, C., Pérez-Ramos, I. M., Marañón, T., Navarro-Fernández, C. M., Olmo, M., et al. (2018). Ett multidimensionellt funktionellt egenskapssätt avslöjar avtrycket av miljöstress i Medelhavets vedartade Communities. Ecosystems 21, 248-262. doi: 10.1007/s10021-017-0147-7
CrossRef Full Text | Google Scholar
Díaz, S., Kattge, J., Cornelissen, J. H., Wright, I. J., Lavorel, S., Dray, S., et al. (2016). Det globala spektrumet av växters form och funktion. Nature 529, 167-171. doi: 10.1038/nature16489
PubMed Abstract | CrossRef Full Text | Google Scholar
Díaz, S., Lavorel, S., de Bello, F., Quétier, F., Grigulis, K. och Robson, T. M. (2007). Integrering av effekter av växternas funktionella mångfald i bedömningar av ekosystemtjänster. Proc. Natl. Acad. Sci. U.S.A 104, 20684-20689. doi: 10.1073/pnas.0704716104
PubMed Abstract | CrossRef Full Text | Google Scholar
Donoghue, M. J., and Edwards, E. J. (2014). Biomskiften och nischutveckling hos växter. Annu. Rev. Ecol. Evol. Syst. 45, 547-572. doi: 10.1146/annurev-ecolsys-120213-091905
CrossRef Full Text | Google Scholar
Douma, J. C., Aerts, R., Witte, J. P. M., Bekker, R. M., Kunzmann, D., Metselaar, K., et al. (2012). En kombination av funktionellt olika växtegenskaper ger ett sätt att kvantitativt förutsäga ett brett spektrum av artsammansättningar i nordvästra Europa. Ecography 35, 364-373. doi: 10.1111/j.1600-0587.2011.07068.x
CrossRef Full Text | Google Scholar
Engemann, K., Sandel, B., Enquist, B. J., Jørgensen, P. M., Kraft, N., Marcuse-Kubitza, A., et al. (2016). Mönster och drivkrafter för dominans av växtfunktionella grupper över det västra halvklotet: en makroekologisk omvärdering baserad på ett massivt botaniskt dataset. Bot. J. Linn. Soc. 180, 141-160. doi: 10.1111/boj.12362
CrossRef Full Text | Google Scholar
Enquist, B. J., Condit, R., Peet, R., Thiers, B. och Schildhauer, M. (2016). Nätverket för botanisk information och ekologi (BIEN): cyberinfrastruktur för ett integrerat botaniskt informationsnätverk för att undersöka de ekologiska effekterna av globala klimatförändringar på växternas biologiska mångfald. PeerJ Prepr. 4:e2615v2. doi: 10.7287/peerj.preprints.2615v2
CrossRef Full Text | Google Scholar
Garnier, E., Cortez, J., Billès, G., Navas, M. L., Roumet, C., Debussche, M., et al. (2004). Växternas funktionella markörer fångar ekosystemegenskaper under sekundär succession. Ecology 85, 2630-2637. doi: 10.1890/03-0799
CrossRef Full Text | Google Scholar
Goldsmith, G. R., Morueta-Holme, N., Sandel, B., Fitz, E. D., Fitz, S. D., Boyle, B., et al. (2016). Plant-O-Matic: en dynamisk och mobil guide till alla växter i Amerika. Methods Ecol. Evol. 7, 960-965. doi: 10.1111/2041-210X.12548
CrossRef Full Text | Google Scholar
Goolsby, E. W., Bruggeman, J., and Ané, C. (2017). Rphylopars: snabba multivariata fylogenetiska komparativa metoder för saknade data och variation inom arten. Methods Ecol. Evol. 8, 22-27. doi: 10.1111/2041-210X.12612
CrossRef Full Text | Google Scholar
Grenié, M., Denelle, P., Tucker, C. M., Munoz, F. och Violle, C. (2017). funrar: an R package to characterize functional rarity. Divers. Distrib. 23, 1365-1371. doi: 10.1111/ddi.12629
CrossRef Full Text | Google Scholar
Hortal, J., de Bello, F., Diniz-Filho, J. A. F., Lewinsohn, T. M., Lobo, J. M. och Ladle, R. J. (2015). Sju brister som hänger samman med storskalig kunskap om biologisk mångfald. Annu. Rev. Ecol. Evol. Syst. 46, 523-549. doi: 10.1146/annurev-ecolsys-112414-054400
CrossRef Full Text | Google Scholar
Kerkhoff, A. J., Enquist, B. J., Elser, J. J., and Fagan, W. F. (2005). Plant allometry, stoichiometry and the temperature-dependence of primary productivity. Glob. Ecol. Biogeogr. 14, 585-598. doi: 10.1111/j.1466-822X.2005.00187.x
CrossRef Full Text | Google Scholar
Kerkhoff, A. J., Fagan, W. F., Elser, J. J. och Enquist, B. J. (2006). Fylogenetisk och tillväxtformsvariation vid skalning av kväve och fosfor i fröväxter. Am. Nat. 168, E103-E122. doi: 10.1086/507879
PubMed Abstract | CrossRef Full Text | Google Scholar
Kraft, N. J., Valencia, R. och Ackerly, D. D. (2008). Funktionella egenskaper och nischbaserad sammansättning av trädsamhällen i en Amazonasskog. Science 322, 580-582. doi: 10.1126/science.1160662
PubMed Abstract | CrossRef Full Text | Google Scholar
Lamanna, C., Blonder, B., Violle, C., Kraft, N. J., Sandel, B., et al. (2014). Funktionellt egenskapsutrymme och den latitudinala mångfaldsgradienten. Proc. Natl. Acad. Sci. U.S.A. 111, 13745-13750. doi: 10.1073/pnas.1317722111
PubMed Abstract | CrossRef Full Text | Google Scholar
Lavorel, S., and Garnier, E. (2002). Prediktion av förändringar i samhällets sammansättning och ekosystemets funktion utifrån växtegenskaper: återigen en ny prövning av den heliga graalen. Funct. Ecol. 16, 545-556. doi: 10.1046/j.1365-2435.2002.00664.x
CrossRef Full Text | Google Scholar
Lavorel, S., Grigulis, K., Lamarque, P., Colace, M.-P., Garden, D., Girel, J., et al. (2011). Användning av funktionella egenskaper hos växter för att förstå landskapsfördelningen av flera ekosystemtjänster. J. Ecol. 99, 135-147. doi: 10.1111/j.1365-2745.2010.01753.x
CrossRef Full Text | Google Scholar
Li, Y., Shipley, B., Price, J. N., Dantas, V. de L., Tamme, R., Westoby, M., et al. (2018). Habitatfiltrering bestämmer den funktionella nischbeläggningen av växtsamhällen över hela världen. J. Ecol. 106, 1001-1009. doi: 10.1111/1365-2745.12802
CrossRef Full Text | Google Scholar
Magallón, S., Gómez-Acevedo, S., Sánchez-Reyes, L. L., and Hernández-Hernández, T. (2015). Ett metakalibrerat tidsträd dokumenterar den tidiga ökningen av blomväxternas fylogenetiska mångfald. New Phytol. 207, 437-453. doi: 10.1111/nph.13264
PubMed Abstract | CrossRef Full Text | Google Scholar
Maitner, B. S., Boyle, B., Casler, N., Condit, R., Donoghue, J., Durán, S. M., et al. (2018). BIEN R-paketet: ett verktyg för att få tillgång till databasen Botanical Information and Ecology Network (BIEN). Methods Ecol. Evol. 9, 373-379. doi: 10.1111/2041-210X.12861
CrossRef Full Text | Google Scholar
Moncrieff, G. R., Bond, W. J., and Higgins, S. I. (2016). Revising the biome concept for understanding and predicting global change impacts. J. Biogeogr. 43, 863-873. doi: 10.1111/jbi.12701
CrossRef Full Text | Google Scholar
Novosolov, M., Rodda, G. H., North, A. C., Butchart, S. H. M., Tallowin, O. J. S., Gainsbury, A. M., et al. (2017). Relationen mellan befolkningstäthet och storlek på avstånd återigen undersökt. Glob. Ecol. Biogeogr. 26, 1088-1097. doi: 10.1111/geb.12617
CrossRef Full Text | Google Scholar
Olson, D. M., Dinerstein, E., Wikramanayake, E. D., Burgess, N. D., Powell, G. V. N., Underwood, E. C., et al. (2001). Terrestrial Ecoregions of the world: a new map of life on earth: a new global map of terrestrial ecoregions provides an innovative tool for conserving biodiversity. Bioscience 51, 933-938.doi: 10.1641/0006-3568(2001)0512.0.CO;2
CrossRef Full Text | Google Scholar
Pennington, R. T., Prado, D. E., and Pendry, C. A. (2000). Neotropiska säsongstorra skogar och kvartära vegetationsförändringar. J. Biogeogr. 27, 261-273. doi: 10.1046/j.1365-2699.2000.00397.x
CrossRef Full Text | Google Scholar
Pennington, R. T., Richardson, J. E., and Lavin, M. (2006). Insikter i den historiska konstruktionen av artrika biom från daterade växtfylogenier, neutral ekologisk teori och fylogenetisk samhällsstruktur. New Phytol. 172, 605-616. doi: 10.1111/j.1469-8137.2006.01902.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Penone, C., Davidson, A. D., Shoemaker, K. T., Di Marco, M., Rondinini, C., Brooks, T. M., et al. (2014). Imputering av saknade uppgifter i dataset med livshistoriska egenskaper: vilket tillvägagångssätt är bäst? Methods Ecol. Evol. 5, 961-970. doi: 10.1111/2041-210X.12232
CrossRef Full Text | Google Scholar
Pillar, V. D., Blanco, C. C., Müller, S. C., Sosinski, E. E., Joner, F. och Duarte, L. D. S. (2013). Funktionell redundans och stabilitet i växtsamhällen. J. Veg. Sci. 24, 963-974. doi: 10.1111/jvs.12047
CrossRef Full Text | Google Scholar
Prado, D. E., and Gibbs, P. E. (1993). Mönster i artfördelningen i de torra säsongsskogarna i Sydamerika. Ann. Mo. Bot. Gard. 80, 902-927. doi: 10.2307/2399937
CrossRef Full Text | Google Scholar
Qian, H., and Sandel, B. (2017). Fylogenetisk struktur hos regionala sammansättningar av angiospermer över latitudinella och klimatiska gradienter i Nordamerika. Glob. Ecol. Biogeogr. 26, 1258-1269. doi: 10.1111/geb.12634
CrossRef Full Text | Google Scholar
Reich, P. B., Wright, I. J., Cavender-Bares, J., Craine, J. M., Oleksyn, J., Westoby, M., et al. (2003). Evolutionen av växters funktionella variation: egenskaper, spektrum och strategier. Int. J. Plant Sci. 164, S143-S164. doi: 10.1086/374368
CrossRef Full Text | Google Scholar
Silva de Miranda, P. L., Oliveira-Filho, A., Pennington, R. T., Neves, D. M., Baker, T. R. och Dexter, K. G. (2018). Användning av trädartsinventeringar för att kartlägga biom och bedöma deras klimatiska överlappningar i tropiskt sydamerikanskt lågland. Glob. Ecol. Biogeogr. 27, 899-912. doi: 10.1111/geb.12749
CrossRef Full Text | Google Scholar
Šímová, I., Violle, C., Kraft, N. J. B., Storch, D., Svenning, J.-C., Boyle, B., et al. (2015). Förskjutningar i egenskapernas medelvärden och varianser i nordamerikanska trädsamlingar: artrikedomsmönster är löst relaterade till det funktionella utrymmet. Ecography 38, 649-658. doi: 10.1111/ecog.00867
CrossRef Full Text | Google Scholar
Šímová, I., Violle, C., Svenning, J.-C., Kattge, J., Engemann, K., Sandel, B., et al. (2018). Rumsliga mönster och klimatsamband för viktiga växtegenskaper i Nya världen skiljer sig åt mellan vedartade och örtartade arter. J. Biogeogr. 45, 895-916. doi: 10.1111/jbi.13171
CrossRef Full Text | Google Scholar
Smith, S. A., and Beaulieu, J. M. (2009). Livshistoria påverkar hastigheterna för klimatnischens utveckling hos blommande växter. Proc. Biol. Sci. 276, 4345-4352. doi: 10.1098/rspb.2009.1176
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, S. A., and Brown, J. W. (2018). Konstruktion av en brett inkluderande fylogeni för fröväxter. Am. J. Bot. 105, 302-314. doi: 10.1002/ajb2.1019
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, S. A., and Donoghue, M. J. (2008). Hastigheten på den molekylära evolutionen är kopplad till livshistorien hos blommande växter. Science 322, 86-89. doi: 10.1126/science.1163197
PubMed Abstract | CrossRef Full Text | Google Scholar
Swenson, N. G. (2014). Phylogenetic imputation of plant functional trait databases. ecography 37, 105-110. doi: 10.1111/j.1600-0587.2013.00528.x
CrossRef Full Text
Swenson, N. G., Enquist, B. J., Pither, J., Kerkhoff, A. J., Boyle, B., Weiser, M. D., et al. (2012). Biogeografi och filtrering av funktionell mångfald av vedartade växter i Nord- och Sydamerika. Glob. Ecol. Biogeogr. 21, 798-808. doi: 10.1111/j.1466-8238.2011.00727.x
CrossRef Full Text | Google Scholar
Swenson, N. G., Weiser, M. D., Mao, L., Araújo, M. B., Diniz-Filho, J. A. F., Kollmann, J., et al. (2017). Fylogeni och förutsägelse av trädfunktionell mångfald över nya kontinentala miljöer. Glob. Ecol. Biogeogr. 26, 553-562. doi: 10.1111/geb.12559
CrossRef Full Text | Google Scholar
Umaña, M. N., Mi, X., Cao, M., Enquist, B. J., Hao, Z., Howe, R., et al. (2017). Rollen av funktionell unikhet och rumslig aggregering för att förklara sällsynthet hos träd. Glob. Ecol. Biogeogr. 26, 777-786. doi: 10.1111/geb.12583
CrossRef Full Text | Google Scholar
van Bodegom, P. M., Douma, J. C., and Verheijen, L. M. (2014). Ett helt egenskapsbaserat tillvägagångssätt för modellering av global vegetationsdistribution. Proc. Natl. Acad. Sci. U.S.A. 111, 13733-13738. doi: 10.1073/pnas.1304551110
PubMed Abstract | CrossRef Full Text | Google Scholar
Violle, C., Reich, P. B., Pacala, S. W., Enquist, B. J. och Kattge, J. (2014). Uppkomsten och löftet av funktionell biogeografi. Proc. Natl. Acad. Sci. U.S.A. 111, 13690-13696. doi: 10.1073/pnas.1415442111
PubMed Abstract | CrossRef Full Text | Google Scholar
Violle, C., Thuiller, W., Mouquet, N., Munoz, F., Kraft, N. J. B., Cadotte, M. W., et al. (2017). Funktionell sällsynthet: ekologin hos outliers. Trends Ecol. Evol. 32, 356-367. doi: 10.1016/j.tree.2017.02.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Walker, B., Kinzig, A., and Langridge, J. (1999). Växtattributdiversitet, motståndskraft och ekosystemfunktion: karaktären och betydelsen av dominerande och mindre arter. Ecosystems 2, 95-113. doi: 10.1007/s100219900062
CrossRef Full Text | Google Scholar
Westoby, M. (1998). Ett strategiskt system för växtekologi för bladhöjd och frö (LHS). Plant Soil 199, 213-227. doi: 10.1023/A:1004327224729
CrossRef Full Text | Google Scholar
Woodward, F. I., Lomas, M. R., and Kelly, C. K. (2004). Globalt klimat och fördelningen av växtbiomarker. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 359, 1465-1476. doi: 10.1098/rstb.2004.1525
PubMed Abstract | CrossRef Full Text | Google Scholar
Wright, I. J., Reich, P. B., Westoby, M., Ackerly, D. D., Baruch, Z., Bongers, F., et al. (2004). Det världsomspännande spektrumet för lövekonomi. Nature 428, 821-827. doi: 10.1038/nature02403
PubMed Abstract | CrossRef Full Text | Google Scholar
Xia, J., Niu, S., Ciais, P., Janssens, I. A., Chen, J., Ammann, C., et al. (2015). Gemensam kontroll av terrestrisk bruttoprimärproduktivitet genom växternas fenologi och fysiologi. Proc. Natl. Acad. Sci. U.S.A. 112, 2788-2793. doi: 10.1073/pnas.1413090112
PubMed Abstract | CrossRef Full Text | Google Scholar
Zanne, A. E., Pearse, W. D., Cornwell, W. K., McGlinn, D. J., Wright, I. J. och Uyeda, J. C. (2018). Funktionell biogeografi hos angiospermer: liv vid extremerna. New Phytol. 218, 1697-1709. doi: 10.1111/nph.15114
PubMed Abstract | CrossRef Full Text | Google Scholar
Zanne, A. E., Tank, D. C., Cornwell, W. K., Eastman, J. M., Smith, S. A., FitzJohn, R. G., et al. (2014). Tre nycklar till strålning av angiospermer till frysmiljöer. Nature 506, 89-92. doi: 10.1038/nature12872
PubMed Abstract | CrossRef Full Text | Google Scholar