Bedömning | Biopsykologi | Jämförande | Kognitiv | Utveckling | Språk | Individuella skillnader | Personlighet | Filosofi | Social |
Metoder | Statistik | Klinisk | Pedagogisk | Industriell | Professionella artiklar | Världspsykologi |
Biologisk:Beteendegenetik – Evolutionär psykologi – Neuroanatomi – Neurokemi – Neuroendokrinologi -Neurovetenskap – Psykoneuroimmunologi – Fysiologisk psykologi – Psykofarmakologi(Index, Översikt)
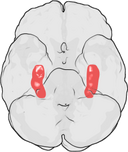
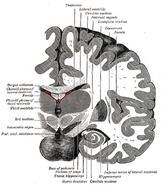
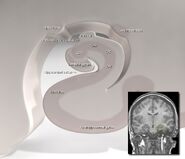
Hippocampus’ placering i den mänskliga hjärnan.
För tidskriften med samma namn, se Hippocampus (tidskrift)
Hippocampus är en del av framhjärnan, belägen i den mediala temporalloben. Den tillhör det limbiska systemet och spelar viktiga roller i långtidsminnet och den rumsliga navigeringen. Människor och andra däggdjur har två hippocampi, en i varje sida av hjärnan. Hos gnagare, där den har studerats mest ingående, är hippocampus formad ungefär som en banan. Hos människor har den en böjd och invecklad form som påminde tidiga anatomer om en sjöhäst. Namnet kommer faktiskt från det grekiska ordet för sjöhäst (grekiska: hippos = häst, campos = hav).
I Alzheimers sjukdom är hippocampus en av de första regionerna i hjärnan som drabbas av skador; minnesproblem och desorientering är några av de första symptomen. Skador på hippocampus kan också uppstå till följd av syrebrist (anoxi), hjärninflammation eller epilepsi i mesiella temporalloben. Personer med omfattande skador på hippocampus kan drabbas av amnesi, det vill säga oförmåga att bilda eller behålla nya minnen.
Hippocampus funktioner
Den kanske tidigaste idén var att hippocampus är involverad i luktsinnet: detta tycks ha föreslagits främst på grund av dess placering i hjärnan, bredvid den luktsinnliga hjärnbarken. Det finns fortfarande ett visst intresse för hippocampus luktreaktioner, men nästan ingen tror nu att hippocampus primära funktion är luktreaktioner.
Under årens lopp har tre huvudidéer om hippocampus funktion dominerat litteraturen: hämning, minne och rymd. Teorin om beteendeinhibition (karikerad av O’Keefe och Nadel som “trampa på bromsen!”) var mycket populär fram till 1960-talet. Den hämtade mycket av sin kraft från två observationer: för det första tenderar djur med hippocampusskador att vara hyperaktiva, för det andra har djur med hippocampusskador ofta svårt att lära sig att hämma reaktioner som de tidigare har lärt sig. Jeffrey Gray utvecklade denna tankegång till en fullfjädrad teori om hippocampus roll i ångest. Inhibitionsteorin är dock inte särskilt populär för närvarande.
Den andra viktiga tankegången relaterar hippocampus till minnet. Även om den hade föregångare fick denna idé sin huvudsakliga kraft från en mycket välkänd rapport av Scoville och Milner om resultaten av kirurgisk förstörelse av hippocampus (i ett försök att lindra epileptiska anfall), hos en patient känd som H.M. Det oväntade resultatet var allvarlig minnesförlust: H.M. kunde inte medvetet minnas händelser som inträffade efter operationen eller under flera år dessförinnan. Fallet väckte ett så stort intresse att H.M. nu sägs vara det mest intensivt studerade medicinska fallet i historien. Under de följande åren har även andra patienter med liknande nivåer av skador på hippocampus och minnesförlust (orsakade av olycka eller sjukdom) studerats, och bokstavligen tusentals experiment har studerat fysiologin bakom neuronal plasticitet i hippocampus. Det råder nu nästan allmän enighet om att hippocampus spelar någon form av viktig roll för minnet, men den exakta karaktären av denna roll är fortfarande mycket omdiskuterad.
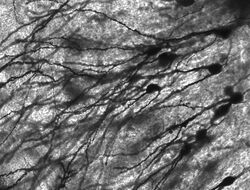
Golgi-färgade neuroner i gnagarnas hippocampus.
Roll i minnet
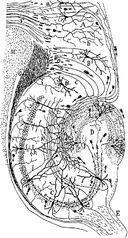
Teckning av de neurala kretsarna i gnagarnas hippocampus. S. Ramón y Cajal, 1911.
Psykologer och neurovetare tvistar om hippocampus exakta roll, men är i allmänhet överens om att den har en väsentlig roll i bildandet av nya minnen om personligt upplevda händelser (episodiskt eller självbiografiskt minne). Vissa forskare föredrar att betrakta hippocampus som en del av ett större minnessystem i den mediala temporalloben som ansvarar för det allmänna deklarativa minnet (minnen som uttryckligen kan verbaliseras – dessa skulle till exempel inkludera minnet av fakta utöver det episodiska minnet).
Huvudartikel: Hippocampus roll i minnet
En del bevis tyder på att hippocampus är inblandad i lagring och bearbetning av rumslig information. Studier på råttor har visat att neuronerna i hippocampus har spatiala skjutfält. Dessa celler kallas för platsceller. Vissa celler avfyras när djuret befinner sig på en viss plats, oavsett färdriktning, medan de flesta är åtminstone delvis känsliga för huvudets riktning och färdriktning.
Huvudartikel: Hippocampus roll i rumsligt minne och navigering
Hippocampus och depression
- Förlust av hippocampusvolym, som kanske återspeglar effekterna av överdrivna glukokortikoider på
neurogenesen, har identifierats i grupper av deprimerade personer i jämförelse med kontrollpersoner.
Huvudartikel: Depression och hippocampus
Historia
Anatomisten Giulio Cesare Aranzi (cirka 1564) använde först termen hippocampus för att beskriva hjärnorganet på grund av dess visuella likhet med en sjöhäst. Detta organ förknippades till en början med luktsinnet, snarare än med dess kända funktion för minnesinhämtning. Ryssen Vladimir Bekhterev noterade hippocampus roll för minnet runt 1900, baserat på observationer av en patient med djupgående minnesstörningar. Under många år var dock den konventionella synen på hippocampus att den, liksom resten av det limbiska systemet, var ansvarig för känslor.
Hippocampus betydelse för minnet uppmärksammades av forskarna av patienten HM. HM led av ett antal anterograda och tidsmässigt graderade retrograda minnesstörningar (sådana störningar är ämnet för filmen Memento) efter det bilaterala avlägsnandet av olika strukturer i den medial-temporala loben (inklusive bilateral ablation av hans hippocampi) för att lindra frekventa epileptiska anfall. Särskilt viktigt är att HM fortfarande kunde lära sig proceduruppgifter (som är förknippade med striatum) och att han hade en IQ som låg över genomsnittet. HM uppvisade en slående singeldissociation mellan intelligens och deklarativt minne. Hippocampusformationens relativa storlek i förhållande till hjärnans totala volym är ofta bevarad hos de flesta däggdjursarter. Trots detta har man funnit att dessa områden är relativt hypotrofa hos valar.
Anatomi
Huvudartikel: Hippocampus anatomi
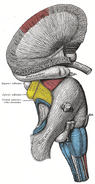
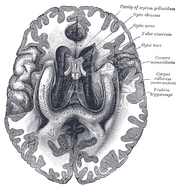
Human hippocampus.
Nissl-färgat koronalt snitt av hjärnan hos en makakakapa, som visar hippocampus (inringad). Källa: brainmaps.org
Anatomiskt sett är hippocampus en utbyggnad av kanten avcortex. Den kan urskiljas som en zon där hjärnbarken smalnar av till ett enda lager av mycket tätt packade neuroner, som krullar sig till en snäv S-form. De strukturer som ligger i kanten av hjärnbarken utgör det så kallade limbiska systemet (latin limbus = gräns): dessa inkluderar hippocampus, cingulate cortex, olfactorycortex och amygdala. Paul MacLean föreslog en gång, inom ramen för sin teori om den tredelade hjärnan, att de limbiska strukturerna utgör den neurala grunden för känslor. De flesta neurovetenskapsmän anser dock inte längre att begreppet “limbiskt system” är giltigt.
Hippocampus, som helhet, ser till slut ut som ett böjt rör, som på olika sätt har analogiserats med en sjöhäst, ett vädjurshorn (Cornu Ammonis) eller en banan.
Denna generella utformning gäller för hela spektrumet av däggdjursarter, från igelkott till människa, även om detaljerna varierar. Hos råttan ser de två hippocampi förbluffande nog ut som ett par bananer som är sammanfogade vid stjälken. I människans och apans hjärnor är den nedre delen av hippocampus, nära tinninglobens bas, mycket bredare än den övre delen. En av konsekvenserna av denna komplexa geometri är att tvärsnitt genom hippocampus kan visa en förbryllande mängd olika former, beroende på snittets vinkel och placering.
Hippocampus har sina starkaste förbindelser med den entorhinala cortexen (EC), som ligger bredvid den i tinningloben. De ytliga lagren av EC ger de mest talrika ingångarna till hippocampus, och de djupa lagren av EC tar emot de mest talrika utgångarna. EC är i sin tur starkt och ömsesidigt kopplad till många andra delar av hjärnbarken. Hippocampus får också en mycket viktig projektion från det mediala septumområdet. Förstörelse av det septala området avskaffar hippocampus thetarytm och försämrar allvarligt vissa typer av minnen. (Så kallade “date rape”-droger anses utöva sina minnesdämpande effekter åtminstone delvis genom att motverka den kolinerga projektionen från det mediala septumområdet till hippocampus.)
Fysiologi
Exempel på råttans hippocampala EEG och CA1 neurala aktivitet i theta (vaken/beteende) och LIA (långsam vågsömn) lägen. Varje diagram visar 20 sekunders data, med ett hippocampalt EEG-spår högst upp, spikraster från 40 samtidigt registrerade CA1-pyramidceller i mitten (varje rasterlinje representerar en annan cell) och ett diagram över löphastighet längst ner. Den översta plotten representerar en tidsperiod under vilken råttan aktivt sökte efter utspridda matpellets. För den nedre plotten sov råttan.
Hippocampus uppvisar två huvudsakliga “lägen” av aktivitet, vart och ett förknippat med ett distinkt mönster av EEG-vågor och aktivitet hos den neurala populationen.Dessa lägen är uppkallade efter de EEG-mönster som är förknippade med dem: theta och stor oregelbunden aktivitet (LIA). Här är några av deras viktigaste egenskaper hos råttan, det djur som har studerats mest ingående:
Theta-läget uppträder under aktiva, alerta beteenden (särskilt rörelse) och även under REM-sömn (drömmar). I thetaläget domineras EEG av stora regelbundna vågor med ett frekvensområde på 6-9 Hz, och huvudgrupperna av hippocampala neuroner (pyramidala celler och granuleceller) uppvisar sparsam populationsaktivitet, vilket innebär att under ett kort tidsintervall är det stora flertalet celler tysta, medan den lilla återstående fraktionen avfyras med relativt hög hastighet, upp till 50 spikar på en sekund för de mest aktiva av dem. En aktiv cell förblir vanligtvis aktiv i mellan en halv sekund och ett par sekunder. När råttan beter sig blir de aktiva cellerna tysta och nya celler blir aktiva, men den totala andelen aktiva celler förblir mer eller mindre konstant. I många situationer bestäms cellaktiviteten till stor del av djurets rumsliga placering, men andra beteendevariabler påverkar den också tydligt.
LIA-läget uppträder under långsam vågsömn (utan att drömma) och även under tillstånd av vaken orörlighet, t.ex. när man vilar eller äter. I LIA-läget domineras EEG av skarpa vågor, som är slumpmässigt tidsbestämda stora avböjningar av EEG-signalen som varar i 200-300 sekunder. Dessa skarpa vågor bestämmer också befolkningens neurala aktivitetsmönster. Mellan dem är pyramidala celler och granuleceller mycket lugna (men inte tysta). Under en skarp våg kan så många som 5-10 % av populationen avge aktionspotentialer under en period av 50 msek; många av dessa celler avger inte en utan en serie spikar.
Dessa två hippocapampala aktivitetslägen kan ses hos både primater och råttor, med det viktiga undantaget att det har varit svårt att se en robust theta-rytmisk aktivitet i hippocampus hos primater. Det finns dock kvalitativt liknande skarpa vågor och liknande statsberoende förändringar i den neurala befolkningsaktiviteten.
Thetarytmen
På grund av sina tätt packade neurala lager genererar hippocampus några av de största EEG-signalerna av alla hjärnstrukturer. I vissa situationer domineras EEG-signalen av regelbundna vågor som ofta pågår i flera sekunder. Detta EEG-mönster är känt som thetarytmen. Det var ett av de tidigaste EEG-fenomenen som upptäcktes: den första beskrivningen kom från Jung och Kornmuller 1938. Det var dock inte förrän 1954, när Green och Arduini publicerade en lång och grundlig studie av thetarytmen hos kaniner, katter och apor, som intresset verkligen tog fart.Kanske till stor del på grund av att de kopplade thetarytmen till upphetsning, som var det heta ämnet för dagen, framkallade deras artikel en flodvåg av uppföljningsstudier, vilket resulterade i att bokstavligen hundratals studier av thetas fysiologi och farmakologi publicerades under 1950- och 1960-talen. Trots detta ganska skrämmande arbete förblev många frågor obesvarade, särskilt frågan om funktion. Även denna mest kritiska fråga har ännu inte fått något övertygande svar.
Teta-rytmicitet är mycket tydlig hos kaniner och gnagare, och förekommer också tydligt hos katter och hundar. Huruvida theta kan ses hos primater är en irriterande fråga. Green och Arduini rapporterade endast mycket korta utbrott av ganska oregelbunden rytmicitet hos apor, och de flesta senare studier har inte sett mycket mer. Variationer i metodik har dock gjort det svårt att dra starka slutsatser.
I råttor (de djur som har studerats i särklass mest ingående) ses theta huvudsakligen under två förhållanden: för det första när djuret går eller på något annat sätt aktivt interagerar med sin omgivning och för det andra under REM-sömn. Frekvensen ökar som en funktion av löphastigheten och börjar vid cirka 6,5 Hz i den lägre delen och ökar till cirka 9 Hz i den högre delen, även om högre frekvenser ibland ses vid dramatiska rörelser, t.ex. vid hopp över stora klyftor. Hos andra, större djurarter är thetafrekvenserna i allmänhet lite lägre. Beteendeberoendet verkar också variera mellan olika arter: hos katter och kaniner observeras thetafrekvenser ofta under tillstånd av orörlig vakenhet. Detta har också rapporterats för råttor, men endast när de är allvarligt skrämda.
Teta är inte bara begränsat till hippocampus. Hos råttor kan den observeras i många delar av hjärnan, inklusive nästan alla som interagerar starkt med hippocampus. Pacemakern för rytmen tros ligga i det mediala septumområdet: detta område projicerar sig till alla de regioner som uppvisar thetarytmicitet, och om det förstörs elimineras theta i hela hjärnan. (Det kan finnas ett undantag, ett litet område i hypotalamus som kallas supramamillarynukleus, som verkar kunna upprätthålla theta oberoende av septum i vissa situationer.)
Thetas funktion, om man antar att den har en sådan, har ännu inte förklarats på ett övertygande sätt, även om många teorier har föreslagits.Den mest populära trenden har varit att koppla den till inlärning och minne.Det är väl etablerat att skador på det mediala septum–den centrala noden i thetasystemet–förorsakar allvarliga störningar av minnet.Det mediala septumet är dock mer än bara kontrollanten av theta,det är också huvudkällan till de kolinerga projektionerna tillhippocampus. Det har inte fastställts att skador på septum har en specifik effekt genom att eliminera theta.
Skarpa vågor
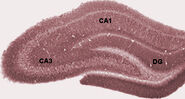
Under sömn, eller under vakna tillstånd när ett djur vilar eller på annat sätt inte är engagerat i sin omgivning, uppvisar hippocampus EEG ett mönster av oregelbundna långsamma vågor, som är något större i amplitud än thetavågorna. Detta mönster avbryts ibland av stora vågor som kallas skarpa vågor. Dessa händelser är förknippade med utbrott av spikaktivitet, som varar 50-100 ms, i pyramidala celler i CA3 och CA1. De är också förknippade med kortvariga högfrekventa EEG-svängningar som kallas “ripples”. Ripples, med frekvenser i intervallet 150-200 Hz hos råttor, kan vanligtvis endast upptäckas av elektroder som antingen är placerade inuti eller mycket nära CA1-cellkroppsskiktet. Elektroder som är placerade var som helst i hippocampus, eller till och med i angränsande hjärnstrukturer, kan däremot ofta uppfatta skarpa vågor som stora långsamma EEG-avvikelser som varar 200-400 ms.
Hos råttor är de skarpa vågorna mest robusta under sömnen, då de uppträder med en genomsnittlig frekvens på cirka 1 per sekund, men i ett mycket oregelbundet tidsmönster. Skarpa vågor förekommer också under inaktiva vakna tillstånd, men de är mindre frekventa då och vanligen mindre. Skarpa vågor har också observerats i människans tinninglob och i apans hippocampus.I apor är skarpa vågor ganska robusta, men de förekommer inte alls lika ofta som hos råttor.
En av de mest intressanta aspekterna av skarpa vågor är att de tycks vara förknippade med minnet. Wilson och McNaughton 1994, och många senare studier, rapporterade att när hippocampala platsceller har överlappande rumsliga eldningsfält (och därför ofta eldar nästan samtidigt), tenderar de att visa korrelerad aktivitet under sömnen som följer på beteendesessionen. Denna förstärkning av korrelationen, allmänt känd som reaktivering, har visat sig vara begränsad huvudsakligen till skarpa vågor. Det har föreslagits att skarpa vågor i själva verket är reaktiveringar av neurala aktivitetsmönster som memorerats under beteendet och som drivs av en förstärkning av de synaptiska förbindelserna inom hippocampus. Denna idé utgör en nyckelkomponent i teorin om “tvåstegsminnen”, som förespråkas av Buzsaki och andra, enligt vilken minnen lagras i hippocampus under beteendet och senare överförs till neocortex under sömnen: skarpa vågor föreslås driva hebbiska synaptiska förändringar i de neocortiska målen för hippocampusutgångarna.
Evolution
Hippocampus har ett generellt sett likartat utseende hos alla däggdjursarter, från basala arter som igelkottet till de mest “avancerade” som människan.Förhållandet mellan hippocampusstorlek och kroppsstorlek ökar i stort sett, och är ungefär dubbelt så stort hos primater som hos igelkottet. Hippocampus tar alltså upp en mycket större volym av den kortikala manteln hos gnagare än hos primater.
Det finns också ett generellt samband mellan hippocampusstorleken och det rumsliga minnet: när jämförelser görs mellan likartade arter tenderar de arter som har en större kapacitet för rumsligt minne att ha större volymer av hippocampus. Detta förhållande gäller även könsskillnader: hos arter där hanar och honor uppvisar starka skillnader i förmåga till rumsligt minne tenderar de också att uppvisa motsvarande skillnader i hippocampusvolym
Inte däggdjursarter har ingen hjärnstruktur som ser ut som däggdjurens hippocampus, men de har en struktur som anses vara homogen med den. Hippocampus är, som påpekats ovan, i huvudsak den mediala kanten av hjärnbarken. Endast däggdjur har en fullt utvecklad hjärnbark, men den struktur som den utvecklats från, som kallas pallium, finns hos alla ryggradsdjur, även de mest primitiva, t.ex. nejonögon eller hagfiskar. Pallium brukar delas in i tre zoner: mediala, laterala och dorsala. Det mediala palliumet utgör föregångaren till hippocampus. Det liknar inte hippocampus visuellt, eftersom lagren inte är s-formade eller omslutna av gyrus dentata, men homologin indikeras av starka kemiska och funktionella släktskap. Det finns nu bevis för att dessa hippocampaliknande strukturer är involverade i den spatiala kognitionen hos fåglar, reptiler och fiskar.
Hos fåglar är överensstämmelsen tillräckligt väletablerad för att de flesta anatomiker hänvisar till den mediala palliala zonen som “avianhippocampus”. Många fågelarter har starka rumsliga färdigheter, särskilt de som gömmer mat. Det finns belägg för att fåglar som gömmer mat har en större hippocampus än andra fågelarter, och att skador på hippocampus orsakar försämringar av det spatiala minnet.
Historien för fiskar är mer komplicerad. Hos teleostfiskar (som utgör den stora majoriteten av de existerande arterna) är framhjärnan konstigt förvrängd i jämförelse med andra typer av ryggradsdjur. De flesta neuroanatomer tror att teleosternas framhjärna i huvudsak är vänd, som en strumpa som vänds ut och in, så att strukturer som hos de flesta ryggradsdjur ligger på insidan, bredvid ventriklarna, finns på utsidan hos teleostfiskar, och tvärtom. En av konsekvenserna av detta är att det mediala pallium (hippocampuszonen) hos ett typiskt ryggradsdjur anses motsvara det laterala palliumet hos en typisk fisk. Flera typer av fiskar (särskilt guldfiskar) har experimentellt visat sig ha ett starkt rumsligt minne och kan till och med bilda “kognitiva kartor” över de områden där de vistas. Det finns bevis för att skador på laterala pallium försämrar det rumsliga minnet (långdistansnavigering, t.ex. laxens hemgång, tycks dock bero på andra mekanismer).
Hippocampusregionens roll i navigeringen tycks alltså börja långt tillbaka i ryggradsdjurens utveckling, före de uppdelningar som ägde rum för flera hundra miljoner år sedan. Man vet ännu inte om det mediala palliumet spelar en liknande roll hos ännu mer primitiva ryggradsdjur, t.ex. hajar och rockor, eller till och med nejonögon och hagfiskar. Vissa typer av insekter och blötdjur som t.ex. bläckfisken har också en stark förmåga till rumslig inlärning och navigering, men dessa verkar fungera på ett annat sätt än det rumsliga systemet hos däggdjur, så det finns ännu ingen god anledning att tro att de har ett gemensamt evolutionärt ursprung.
Se även
- Amygdalohippocampectomy
- Median forebrain bundle
- Septal nuclei
Referenser & Bibliografi
Nyckeltexter
Böcker
- Aboitiz, F, Morales D, Montiel J (2003). Det evolutionära ursprunget till däggdjurens isocortex: Mot ett integrerat utvecklingsmässigt och funktionellt tillvägagångssätt. Behav. Brain Sciences 26: 535-552.
- Amaral, D; Lavenex P (2006). “Ch 3. Hippocampal Neuroanatomy” Andersen P, Morris R, Amaral D, Bliss T, O’Keefe J The Hippocampus Book, Oxford University Press.
- Broglio, C, Gómez A, Durán E, Ocaña FM, Jiménez-Moya F, Rodríguez F, Salas C (2002). Kännetecken för en gemensam plan för vertebraternas framhjärna: Specialiserade palliala områden för rumsligt, tidsmässigt och känslomässigt minne hos aktinopterygiska fiskar. Brain Res. Bull. 57: 397-399.
- Buzsáki, G (2002). Theta-svängningar i hippocampus. Neuron 33: 325-340.
- Buzsáki, G (2006). Rhythms of the Brain.
- Cantero, JL, Atienza M, Stickgold R, Kahana MJ, Madsen JR, Kocsis B (2003). Sömnberoende theta-svängningar i människans hippocampus och neocortex. J. Neurosci. 23: 10897-10903.
- Colombo, M, Broadbent N (2000). Är fågelhippocampus en funktionell homolog till däggdjurens hippocampus. Neurosci. Biobehav. Rev. 24: 465-484.
- Eichnbaum, H; Cohen NJ (1993). Memory, Amnesia, and the Hippocampal System, MIT Press.
- Ekstrom, AD, Kahana MJ, Caplan JB, Fields TA, Isham EA, Newman EL, Fried I (2003). Cellulära nätverk som ligger till grund för människans rumsliga navigering. Nature 425: 184-188.
- Gray, JA; McNaughton N (2000). Neuropsykologi för ångest: An Enquiry into the Functions of the Septo-Hippocampal System, Oxford University Press.
- Green, JD, Arduini AA (1954). Hippocampal electric activity in arousal. J. Neurophysiol. 17: 533-557.
- Jacobs, LF, Gaulin SJ, Sherry DF, Hoffman GE (1990). Evolution av rumslig kognition: könsspecifika mönster av rumsligt beteende förutsäger hippocampal storlek. PNAS 87: 6349-6352.
- Jacobs, LF (2003). Utvecklingen av den kognitiva kartan. Brain Behav. Evol. 62: 128-139.
- Kirk, IJ, McNaughton N (1991). Supramammillär celleldning och hippocampal rytmisk långsam aktivitet. Neuroreport 11: 723-725.
- Maguire, EA, Gadian DG, Johnsrude IS, Good CD, Ashburner J, Frackowiak RS, Frith CD (2000). Navigationsrelaterade strukturella förändringar i hippocampi hos taxichaufförer. PNAS 97: 4398-4403.
- McNaughton, BL, Battaglia FP, Jensen O, Moser EI, Moser MB (2006). Path integration and the neural basis of the ‘cognitive map’. Nat. Rev. Neurosci. 7: 663-678.
- Moser, EI, Kropf E, Moser M-B (2008). Place Cells, Grid Cells, and the Brain’s Spatial Representation System. Ann. Rev. Neurosci. 31.
- Nieuwenhuys, R (1982). En översikt över organisationen av hjärnan hos aktinopterygiska fiskar. Am. Zool. 22: 287-310.
- O’Kane, G, Kensinger EA, Corkin S (2004). Bevis för semantisk inlärning vid djup amnesi: En undersökning med patienten H.M.. Hippocampus 14: 417-425.
- O’Keefe, J; Nadel L (1978). The Hippocampus as a Cognitive Map, Oxford University Press.
- Portavella, M, Vargas JP, Torres B, Salas C (2002). Effekterna av telencefaliska palliella lesioner på rumslig, temporal och emotionell inlärning hos guldfiskar. Brain Res. Bull. 57: 397-399.
- Rodríguez, F, Lópeza JC, Vargasa JP, Broglioa C, Gómeza Y, Salas C (2002). Rumsligt minne och hippocampal pallium genom vertebraternas evolution: insikter från reptiler och teleostfiskar. Brain Res. Bull. 57: 499-503.
- Sainsbury, RS, Heynen A, Montoya CP (1987). Beteendemässiga korrelat av hippocampal typ 2 theta hos råtta. Physiol. Behav. 39: 513-519.
- Scoville, WB, Milner B (1957). Förlust av nyligt minne efter bilaterala hippocampala lesioner. J. Neurol. Neurosurg. Psych. 20: 11-21.
- Shettleworth, SJ (2003). Memory and Hippocampal Specialization in Food-Storing Birds: Challenges for Research on Comparative Cognition. Brain Behav. Evol. 62: 108-116.
- Skaggs, WE, McNaughton BL, Wilson MA, Barnes CA (1996). Theta fas precession i hippocampala neuronala populationer och komprimering av temporala sekvenser. Hippocampus 6: 149-176.
- Squire, LR (1992). Minnet och hippocampus: en sammanfattning av resultat från rötter, apor och människor. Psych. Rev. 99: 195-231.
- Squire, LR; Schacter DL (2002). The Neuropsychology of Memory, Guilford Press.
- Vanderwolf, CH (1969). Hippocampal elektrisk aktivitet och frivillig rörelse hos råttan. EEG & Clin. Neurophysiol. 26: 407-418.
- Vargas, JP, Bingman VP, Portavella M, López JC (2006). Telencephalon och geometriskt utrymme hos guldfiskar. Eur. J. Neurosci. 24: 2870-2878.
- West, MJ (1990). Stereologiska studier av hippocampus: en jämförelse av de hippocampala underavdelningarna hos olika arter inklusive igelkottar, laboratoriegnagare, vilda möss och människor…. Prog. Brain Res. 83: 13-36.
Papers
- Amaral DG och Cowan WM. 1980. Subkortikala afferenter till hippocampusformationen hos apan. Journal of Comparative Neurology. Feb 15; 189(4):573-91.
- Duvernoy, H. (2005) The Human Hippocampus, 3rd ed. Berlin: Springer-Verlag.
Ytterligare material
Papper
- Google Scholar
- http://www.psycheducation.org/emotion/hippocampus.htm Stora hjärnturer!!!
- BrainMaps at UCDavis hippocampus
- Artificial Hippocampus
- BrainInfo at the University of Washington hier-164
- John O’Keefe & Lynn Nadel (1978) The Hippocampus as a Cognitive Map , Oxford University Press. Fulltext
Ytterligare bilder
Tillägg ett foto till detta galleri
Humana hjärnan: Limbiskt system
Amygdala – Cingulate gyrus – Fornicate gyrus – Hippocampus – Hypothalamus – Mammillary body – Nucleus accumbens – Orbitofrontal cortex – Parahippocampal gyrus
|
v-d-e
Människans hjärna: Förhjärna (cerebrum – hjärnbark – hjärnhalvor, grå substans) (TA A14.1.09.002-240, 301-320, GA 9.818-826) |
|||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Frontalloben |
|
||||||||||||||||
| Parietalloben |
|
Superolaterala
|
Medial/inferior
|
Både
|
|||||||||||||
| Occipitalloben |
|
Superolateralt
|
Medial/inferior
|
||||||||||||||
| Temporalloben |
|
Superolateralt
|
Medial/inferior
|
||||||||||||||
| Interlobar sulci/fissurer |
|
Superolateralt
|
Medial/inferior
|
||||||||||||||
| Limbic lobe |
|
Andra
|
|||||||||||||||
| Insulära loben |
Lång gyrus i insula – Kort gyri i insula – Cirkulär sulcus i insula |
||||||||||||||||
| Allmänt |
Operculum – Polerna i hjärnhalvorna |
||||||||||||||||
Vissa kategoriseringar är approximationer, och vissa Brodmann-områden sträcker sig över gyri.
|
|||||||||||||||||
Denna sida använder innehåll med Creative Commons-licens från Wikipedia (se författare).