- Introduction
- Methodes
- Distributiegegevens en bioomclassificatie
- Species Composition Among Biomes
- Trait Data
- Trait Hypervolumes Meting
- Functioneel onderscheidend vermogen en wijdverspreidheid
- Gelijkenis tussen functionele ruimte en biome
- Discussie
- Kennislacunes
- Gemeenschappelijke patronen binnen biomen
- Vergelijkingen tussen biomen
- Conclusies
- Beschikbaarheid van gegevens
- Bijdragen van auteurs
- Funding
- Conflict of Interest Statement
- Acknowledgments
- Aanvullend materiaal
Introduction
Ecologen en biogeografen ordenen de biodiversiteit van landplanten in klimatologisch bepaalde biomen, met fysiognomieën die worden gekenmerkt door de groeivormen en functionele kenmerken van de dominante soorten (Moncrieff et al., 2016). Het concept van de biomen heeft inderdaad een lange geschiedenis die teruggaat tot Carl Ludwig Willdenow en Alexander von Humboldt. Willdenow erkende dat vergelijkbare klimaten vergelijkbare vegetatievormen ondersteunen, en Humboldt observeerde het wijdverbreide verband tussen plantendistributie, fysionomie en omgevingsfactoren. In het algemeen weerspiegelt het concept van de bioom de veronderstelling dat gelijksoortige milieudruk selecteert op soorten met gelijksoortige functionele eigenschappen, onafhankelijk van hun evolutionaire geschiedenis. Tegelijkertijd zijn de bestaande biomen op aarde tot op zekere hoogte fylogenetisch verschillend, waarbij veel/meeste van de karakteristieke soorten afkomstig zijn van specifieke lijnen die belangrijke aanpassingen vertonen, niet alleen aan klimaatselectie maar ook aan bijkomende druk zoals vuur of megaherbivoren (Woodward et al., 2004; Pennington et al., 2006; Donoghue en Edwards, 2014). Omdat biomen breedschalige regelmatigheden in de vegetatie van de aarde vertegenwoordigen, is het begrijpen van functionele verschillen tussen biomen van cruciaal belang voor het modelleren van de mondiale koolstofcyclus en het functioneren van het systeem aarde, met inbegrip van reacties op antropogene mondiale verandering (Bonan et al, 2012; van Bodegom et al., 2014; Xia et al., 2015).
De afgelopen twee decennia is een snelle groei te zien geweest in het begrijpen van de functionele dimensie van plantenbiodiversiteit over continentale schalen (Swenson et al., 2012; Lamanna et al., 2014; Šímová et al., 2018). Uitgebreide inspanningen op het gebied van gegevenssynthese en -analyse hebben een beperkte functionele kenmerkenruimte gedefinieerd waarin variatie in totale plantgrootte, bladeconomie en hydraulica is opgenomen, bekend als functionele kenmerkenspectra (Wright et al., 2004; Díaz et al., 2016). Functionele eigenschap spectra definiëren fysiologische en ecologische trade-offs die de life-history strategieën van planten bepalen (Westoby, 1998; Reich et al., 2003; Craine, 2009) en hun invloed op gemeenschapsvorming (Kraft et al., 2008), ecosysteem functie (Lavorel en Garnier, 2002; Garnier et al., 2004; Kerkhoff et al., 2005; Cornwell et al., 2008), en zelfs de snelheid van evolutie (Smith en Donoghue, 2008; Smith en Beaulieu, 2009). De generaliteit en het nut van functionele kenmerken van planten hebben ertoe geleid dat ze snel zijn opgenomen in modellen van de mondiale verspreiding van vegetatie (van Bodegom et al., 2014), biogeochemische cycli (Bonan et al., 2012), en ecosysteemdiensten (Díaz et al., 2007; Cadotte et al., 2011; Lavorel et al., 2011). Er zijn echter maar weinig studies die uitgebreide metingen van functionele diversiteit op de schaal van het biome hebben onderzocht, over zowel dominante als ondergeschikte soorten en levensvormen.
Er zijn twee contrasterende sets van voorspellingen over hoe functionele diversiteit varieert tussen klimatologisch en fysiognomisch verschillende biomen. Enerzijds, als de biodiversiteit van biomen een afspiegeling is van de variatie in de beschikbare ecologische niche-ruimte, dan zouden minder taxonomisch diverse biomen een kleinere, grotendeels geneste subset moeten vertegenwoordigen van de functionele ruimte die wordt ingenomen door meer diverse biomen. Aan de andere kant, als gevolg van de mondiale aard van fundamentele afwegingen in plantenstructuur en -functie, en vergelijkbare selectiedruk die gelijksoortig werkt tussen soorten, kunnen assemblages die verschillende omgevingen bewonen in feite vergelijkbare gebieden van de karakteristieke ruimte delen (Reich et al., 2003; Wright et al., 2004; Díaz et al., 2016). Recente studies bevestigen dat de relaties tussen klimaat en functionele en taxonomische diversiteit (d.w.z. soortenrijkdom) complex en schaalafhankelijk zijn. In sommige gevallen volgt functionele diversiteit nauw de klimatologische gradiënten, met lagere functionele diversiteit in meer variabele en extreme omstandigheden (Swenson et al., 2012; de la Riva et al., 2018), en het volume van de functionele ruimte in lokale assemblages breidt zich uit met toenemende taxonomische rijkdom (Lamanna et al., 2014; Li et al., 2018). Analyses van soortenpools op regionale schaal suggereren echter dat grote veranderingen in soortenrijkdom gepaard kunnen gaan met minimale effecten op de functionele diversiteit (Lamanna et al., 2014; Šímová et al., 2015). Bovendien kunnen de reacties van taxonomische en functionele diversiteit op het klimaat verschillen tussen de belangrijkste plantengroeivormen (Šímová et al., 2018), en de relatieve diversiteit van verschillende groeivormen kan aanzienlijk veranderen langs de klimaatgradiënten die verschillende biomen definiëren (Engemann et al., 2016).
De vooruitgang in het analyseren van functionele diversiteit op continentale tot mondiale schalen is gedeeltelijk beperkt door gegevenstekorten bij het catalogiseren van de taxonomische, distributieve, fylogenetische en functionele aspecten van biodiversiteit (Hortal et al., 2015). Bovendien zijn er, zelfs gezien de beperkte beschikbare gegevens, aanzienlijke informatica-uitdagingen verbonden aan het standaardiseren en integreren van grote, ongelijksoortige datasets die de geografische distributies, functionele kenmerken en fylogenetische relaties van soorten beschrijven (Violle et al., 2014). Hier onderzoeken we de verdeling van functionele diversiteit van plantensoorten over de biomen van Noord- en Zuid-Amerika met behulp van de Botanical Information and Ecology Network-database (BIEN; Enquist et al., 2016; Maitner et al., 2018), die informatie over de verspreiding en functionele kenmerken van >100.000 soorten landplanten samenvoegt. Specifiek onderzoeken we variatie in de functionele diversiteit en het onderscheidend vermogen van biomen, op basis van verspreidingskaarten van landplantensoorten op een schaal van 100 × 100 km gridcellen en een uitgebreide dataset van zes functionele kenmerken die de belangrijkste assen van variatie in ecologische strategieën weerspiegelen.
Onze doelen in deze studie zijn 3-voudig. Ten eerste, om hardnekkige tekorten aan gegevens aan het licht te brengen, documenteren we de omvang van de beschikbare gegevens die de functionele diversiteit en het onderscheidend vermogen van biomen karakteriseren. Ten tweede, gegeven de beschikbare gegevens, karakteriseren we de verdeling van de functionele diversiteit binnen biomen over zowel dominante als ondergeschikte groeivormen. Deze analyses stellen ons in staat om het functionele onderscheidend vermogen van een bioma beter te kwantificeren door de meest voorkomende functionele strategieën van de meest verspreide soorten binnen het bioma te identificeren. Ten derde vragen we ons af of biomen in feite worden gekenmerkt door functioneel verschillende verzamelingen van soorten, gebaseerd op de overlap van multidimensionale hypervolumes in functionele eigenschappenruimte.
Methodes
Distributiegegevens en bioomclassificatie
Om de effecten van steekproefbias te verminderen die kenmerkend zijn voor voorkomensdatasets samengesteld uit meerdere bronnen, gebruikten we de BIEN 2.0 verspreidingskaarten voor 88.417 van plantensoorten verspreid in Noord- en Zuid-Amerika (Goldsmith et al., 2016). De BIEN-database integreert gestandaardiseerde plantenwaarnemingen afkomstig van herbariumspecimens en vegetatieplotinventarisaties. Kaarten van het verspreidingsgebied van soorten in de BIEN-databases werden geschat met behulp van een van de drie benaderingen, afhankelijk van de beschikbare waarnemingen van elke soort. Voor soorten met slechts één of twee waarnemingen (ca. 35% van de soorten) werd het geografische verspreidingsgebied gedefinieerd als een vierkant gebied van 75.000 km2 rond elk gegevenspunt. De geografische verspreidingsgebieden van soorten met drie of vier waarnemingen zijn vastgesteld met behulp van een convexe romp (ongeveer 15% van de soorten). Ten slotte werden de verspreidingskaarten voor soorten met ten minste vijf voorkomens verkregen met behulp van het Maxent-algoritme voor het modelleren van de verspreiding van soorten, waarbij 19 klimaatlagen als voorspellende variabelen werden gebruikt en 19 ruimtelijke eigenvectoren als filters om de voorspellingen van de modellen te beperken (zie Goldsmith et al, 2016 voor meer details over de methodologie van de verspreidingskaart).
We overlapten de BIEN 2.0 plantensoortenkaarten op een 100 × 100 km rasterkaart met een Lambert Azimuthal Equal Area-projectie om een aanwezigheids-/afwezigheidsmatrix van soorten voor elke rastercel te verkrijgen. Op basis van de bioomclassificatie van Olson et al. (2001) hebben we elke rastercel in de matrix ingedeeld in een van de categorieën van biomen. Vanwege computationele beperkingen voor de daaropvolgende analyses voegden wij sommige biomen samen op basis van hun overeenkomsten in klimaat en vegetatie en literatuur om een brede classificatie te verkrijgen zoals beschreven in tabel 1 (met uitzondering van Inland Water, Rock and Ice en Mangroves). De ecoregio’s Chaco en Caatinga werden geclassificeerd als respectievelijk Xeric Woodlands en Dry forest, in navolging van Prado en Gibbs (1993), Pennington et al. (2000), Banda et al. (2016), Silva de Miranda et al. (2018).
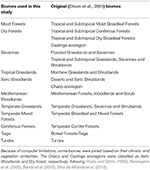
Tabel 1. Overzicht van de in deze studie aangenomen bioomclassificatie en de equivalente (Olson et al., 2001) biomenclassificatie (exclusief Inland Water, Rock and Ice, en Mangroves).
Species Composition Among Biomes
Om de variatie in functionele eigenschappenruimte tussen biomen te begrijpen, onderzochten we eerst de verschillen in plantensoortensamenstelling tussen de biomen. Gebaseerd op de soortenlijst voor elke rastercel en biome, definieerden we de karakteristieke soorten van elk biome als die soorten die het grootste deel van hun verspreidingsgebied in dat biome hebben. Deze lijsten van geografisch overheersende soorten in een bepaald biome werden vergeleken met de lijst van soorten in andere biomes. Deze paarsgewijze vergelijking biedt een eenvoudige manier om een directionele taxonomische overlap tussen biomen af te leiden (d.w.z., het aandeel van de overheersende soorten van een biome dat wordt gedeeld met een ander biome). Van de 88.417 soorten met beschikbare verspreidingskaarten hebben 44.899 soorten een verspreidingsgebied dat meer dan één biome omvat, en 43.518 soorten zijn endemisch voor een specifiek biome.
Trait Data
We hebben alle trait informatie voor plantensoorten in de Nieuwe Wereld die beschikbaar is in de BIEN 3.0 dataset (opgehaald op 7 februari 2018) geëxtraheerd, wat een totaal geeft van 80.405 trait waarnemingen op soortniveau. Vervolgens filterden we de informatie voor zes functionele kenmerken: maximale planthoogte (m), zaadmassa (mg), houtdichtheid (mg/cm3), specifiek-bladoppervlak SLA (cm2/g) en bladfosfor- en bladstikstofconcentratie per massa-eenheid (mg/g). Na filtering bleven er in totaal 18.192 waarnemingen op soortniveau over. Hiervan werd de subset van 8.820 soorten met zowel verspreidingskaarten als kenmerkeninformatie gebruikt voor verdere analyses.
Om de functionele kenmerkenruimte voor elk biome te schatten, hadden we volledige kenmerkengegevens nodig. Daarom imputeerden we ontbrekende traitgegevens fylogenetisch (Bruggeman et al., 2009; Penone et al., 2014; Swenson, 2014; Swenson et al., 2017) met behulp van het R-pakket “Rphylopars” v 0.2.9 (Goolsby et al., 2017) en de recent gepubliceerde fylogenie van zaadplanten door Smith en Brown (2018) als basislijn. Fylogenetische imputatie is een hulpmiddel voor het voorspellen van ontbrekende gegevens in datasets van functionele kenmerken, gebaseerd op de aanname dat nauw verwante soorten de neiging hebben vergelijkbare kenmerkwaarden te hebben (Swenson, 2014). We gebruikten de ALLBM-boom (d.w.z. genenbank en Open Tree of life taxa met een ruggengraat geleverd door Magallón et al., 2015) omdat deze de overlap maximaliseerde tussen soorten met beschikbare trait- en distributie-informatie. Na de trait imputatie bleven in totaal 7.842 soorten met volledige trait informatie en verspreidingskaarten over voor verdere analyse.
Trait Hypervolumes Meting
We hebben de relatieve functionele diversiteit van biomen gemeten door trait hypervolumes te berekenen van soortenpools binnen biomen cellen. Als gevolg van computationele beperkingen, hebben we geconstrueerd de eigenschap hypervolumes met behulp van een aselecte steekproef van 20% van de cellen in elk biome. De hypervolumes voor elke rastercel werden geschat met behulp van de geëxtraheerde zes functionele kenmerken en het R-pakket “hypervolume” (Blonder et al., 2014, 2018), met behulp van de Gaussische KDE-methode met de standaard Silverman bandbreedteschatter. Zaadmassa, hoogte en houtdichtheid werden log-getransformeerd, terwijl SLA kwadratisch-root getransformeerd werd. Alle kenmerken werden vóór de analyse geschaald en gecentreerd. Hypervolumes worden gerapporteerd in eenheden van standaarddeviaties (sd) tot de macht aan het aantal gebruikte kenmerken (d.w.z., sd6).
Functioneel onderscheidend vermogen en wijdverspreidheid
Omdat biomen worden gekenmerkt door hun dominante vegetatie, onderzochten we ook de geografische algemeenheid en het functionele onderscheidend vermogen van soorten binnen biomen. Met behulp van verspreidingskaarten van soorten en informatie over functionele kenmerken, schatten we het functionele onderscheidend vermogen en de wijdverspreidheid van elke soort in elk biome, volgens het conceptuele kader van functionele zeldzaamheid van Violle et al. (2017). Met behulp van de hele set van (gemeten en geïmputeerde) kenmerken hebben we eerst de Euclidische afstand in gestandaardiseerde kenmerkenruimte tussen alle paar soorten gemeten. Vervolgens berekenden we het functionele onderscheidend vermogen (Di) voor elke soort in elk biome als de gemiddelde functionele afstand van een soort tot de N andere soorten binnen de soortenpool van het biome. Di werd geschaald tussen 0 en 1 binnen elk biome, waarbij lagere waarden staan voor soorten die functioneel gemeenschappelijk zijn (redundant) en hogere waarden voor soorten die functioneel onderscheidend zijn, vergeleken met de andere soorten in het biome. Wij schatten ook de geografische wijdverspreidheid (Wi) van een soort in een biome, gemeten als het aantal gridcellen dat door de soort binnen een biome wordt bezet ten opzichte van het totale aantal cellen in dat biome. Een waarde van 1 geeft aan dat de doelsoort aanwezig is in alle gridcellen van het biome. Zowel functioneel onderscheidend vermogen als geografische wijdverspreidheid werden berekend met behulp van het R-pakket “funrar” (Grenié et al., 2017).
Omdat het ons ontbreekt aan uitgebreide maten voor dominantie op basis van lokale abundantie of biomassa, gebruikten we de maten Di en Wi om voor elk biome de meest “gewone” soorten te identificeren als soorten die zowel geografisch wijdverspreid zijn (Wi > 0,5) als functioneel vergelijkbaar (Di < 0,25). Deze laatste drempelwaarde werd gebruikt omdat het derde kwantiel van functionele onderscheidingswaarden tussen biomen varieerde van 0,2 tot 0,3. Om functioneel verschillende soorten te onderscheiden van functioneel gelijkende soorten in de volgende analyses, gebruikten wij daarom een waarde van 0.25 als cut-off (d.w.z, soorten met onderscheidingswaarden < 0.25 werden beschouwd als functioneel gemeenschappelijk of overbodig binnen een specifiek biome).
Gelijkenis tussen functionele ruimte en biome
Om de overlap tussen de hypervolumes van de biomen te schatten, gebruikten we de Sørensen gelijksoortigheidsindex, gebruik makend van (i) het totaal aantal soorten en (ii) de lijst van soorten die beschouwd werden als functioneel gemeenschappelijk en geografisch wijdverspreid voor elk biome. De gelijkenis tussen functionele kenmerken werd berekend als de paarsgewijze fractionele overlap van de hypervolumes tussen de biomen. De fractionele overlapping werd berekend door tweemaal het volume van de intersectie van twee hypervolumes te delen door het volume van hun unies. Alle hypervolumes werden geschat met behulp van het R-pakket “hypervolume” (Blonder et al.We vonden een aanzienlijke overlap in de samenstelling van de plantensoorten in de verschillende biomen, gebaseerd op de kruising van de gemodelleerde soortenbereiken (zie figuur 1, en aanvullende tabel 1). Deze taxonomische overlap is groter binnen tropische en gematigde biomen, met relatief weinig soorten die worden gedeeld tussen deze twee klimatologische zones. Interessant is dat xerische bosgebieden soorten delen met zowel tropische als gematigde biomen. Ondanks het hoge percentage soorten dat kenmerkend is voor tropisch vochtig bos (83%), deelt dit biome ook grote aantallen soorten met andere tropische biomes zoals droge bossen, savannes, xerisch bos en tropische graslanden. De grote taxonomische overlapping van gematigde biomen zoals gematigde graslanden, mediterrane bossen, taiga’s en toendra’s suggereert het potentieel voor een laag functioneel onderscheidend vermogen, waarbij sommige gematigde biomen een armere subgroep vormen van de meer soortenrijke, functioneel diverse tropische biomen.
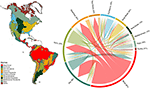
Figuur 1. Overlapping van plantensoorten tussen de biomen van de Nieuwe Wereld. De procentuele waarden geven de fractie weer van de soorten die in een biome voorkomen en die het grootste deel van hun geografische verspreidingsgebied in dat biome hebben, N = 84.413. De basis van elke vertakte pijl is gepositioneerd om het biome aan te geven dat het grootste deel van het bereik van een soort omvat, terwijl de breedte het aantal soorten weergeeft dat wordt gedeeld met de biomes aan de uiteinden van de pijl. Zie aanvullende tabel 1 voor de onderliggende matrix.
Met minder dan 10% van de in kaart gebrachte soorten vertegenwoordigd, de kenmerken gegevens waren vrij schaars voor onze bioom-schaal soortenassemblages (Figuur 2A). Bovendien varieerden de beschikbare kenmerkgegevens aanzienlijk tussen kenmerken, plantenclades en biomen. Fylogenetisch waren vooral blad P en hout dichtheid slecht vertegenwoordigd met hele clades waar geen gegevens beschikbaar waren. Het gevolg was dat van de ongeveer 2/3 van de kenmerkwaarden voor onze 7.842 soorten die moesten worden geïmputeerd, vele moesten worden toegewezen in afwezigheid van nauw verwante taxa. Geografisch gezien zijn SLA, hoogte en zaadmassa onderbemonsterd in de tropen en het gematigde Zuid-Amerika, terwijl blad-N en houtdichtheid gelijkmatiger over de regio’s zijn bemonsterd. Blad P is slecht bemonsterd in alle biomen. Trait hypervolumes gecreëerd uit soortenpools van een willekeurige selectie van 20% van de cellen voor elk biome vertonen vooral verschillen tussen tropische en gematigde/koude biomen (Figuur 2B). Hypervolumes van kenmerken hebben de neiging groter en variabeler te zijn in tropische biomen, met de grootste variatie in vochtige en droge tropische bossen. Bij de gematigde/koude biomen hebben naaldbossen en gematigde gemengde bossen de grootste kenmerk-hypervolumes. Interessant is dat xerische struikgewassen een verdeling van hypervolumes vertonen die meer lijkt op die van tropische dan van gematigde biomen.
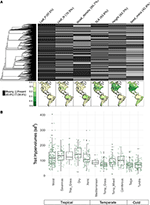
Figuur 2. (A) Percentage soorten in de BIEN 3.0-database met bekende (grijs) of ontbrekende (zwart) kenmerkwaarden. De fylogenie links komt overeen met de zaadplanten ALLBM-boom van Smith en Brown (2018) (d.w.z. GenBank en Open Tree of life taxa met een ruggengraat geleverd door Magallón et al., 2015). Kaarten geven het aandeel weer van soorten met trait informatie ten opzichte van het totale aantal soorten in de BIEN 2.0 database. (B) Verdeling van trait hypervolumes van 20% van willekeurig geselecteerde 100 × 100 km cellen in elk biome. Hypervolumes worden gerapporteerd in eenheden van standaardafwijkingen tot de macht van het aantal gebruikte kenmerken.
De relatie tussen functioneel onderscheidend vermogen en geografische spreiding is opmerkelijk vergelijkbaar tussen biomen (figuur 3). In elk biome is de overgrote meerderheid van de soorten zowel geografisch beperkt als functioneel gelijkaardig. Tegelijkertijd waren de soorten met de meest onderscheidende functies binnen elk biome over het algemeen ook geografisch beperkt, en de soorten met de grootste geografische verspreiding waren bijna altijd functioneel vergelijkbaar.
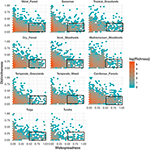
Figuur 3. Patronen van functioneel onderscheidend vermogen in de verschillende biomen. Onderscheidend vermogen geeft aan hoe soorten functioneel van elkaar verwijderd zijn binnen een biome (d.w.z. de gemiddelde paarsgewijze fenotypische afstand van een focale soort tot alle andere soorten). Hoe groter de waarde, hoe verder een soort verwijderd is van het middelpunt van de functionele ruimte van het biome. Wijdverspreidheid meet hoe algemeen een soort geografisch gezien is binnen een biome. Een waarde van 0 geeft aan dat een soort slechts in één enkele biome cel voorkomt. Bin-waarden zijn het aantal soorten op een loge-schaal. Rechthoeken geven de soorten weer die als functioneel gelijkend en wijdverspreid worden beschouwd in elk biome, gebruik makend van afkapwaarden van 0.25 en 0.5 van onderscheidend vermogen en wijdverspreidheid, respectievelijk.
We vonden aanzienlijke variatie in de proportionele verdeling van groeivormen binnen en tussen biomen. In tropische biomen zijn houtachtige soorten de dominante groeivormen in termen van soortenaantallen, terwijl in gematigde milieus kruidachtige soorten domineren (Figuur 4). Wanneer we alleen kijken naar de soorten die wijdverbreid zijn en functioneel veel voorkomen in elk biome, verandert de verdeling van de groeivormen over de biomen, vooral in het aandeel bomen, kruiden en grassen. In tropische biomen bijvoorbeeld, daalde het aandeel bomen in elk biome, behalve in vochtig bos. In gematigde biomen nam het aandeel van grassen toe, vooral in de poolstreken. Interessant is dat de verdeling van groeivormen in xerische bossen meer lijkt op de verdeling in gematigde biomen wanneer alleen wordt gekeken naar wijdverspreide en functioneel veel voorkomende soorten.
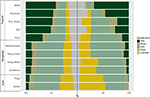
Figuur 4. Distributie van groeivormen in elk biome (links) met behulp van het totale aantal soorten, en (rechts) met behulp van alleen die soorten die functioneel vergelijkbaar en wijdverbreid zijn binnen het biome (zie figuur 3).
Paargewijze vergelijkingen van de soortensamenstelling tussen biomen onthullen drie belangrijke clusters die de tropische, gematigde en polaire klimaatzones vertegenwoordigen (figuur 5A), als gevolg van de grote taxonomische overlap binnen, maar niet tussen, deze regio’s (figuur 1). Paarsgewijze vergelijkingen van kenmerk-hypervolumes tussen biomen laten een minder duidelijke clustering van klimaatzones zien (figuur 5B). In dit geval overlapt de functionele ruimte van xerische bosgebieden aanzienlijk met gematigde biomen. Deze analyse onthult ook de overlapping in functionele ruimte van taiga’s met gematigde graslanden en gemengde bossen. De paarsgewijze vergelijking van kenmerk-hypervolumes tussen biomen, waarbij alleen die soorten worden gebruikt die als functioneel gewoon en wijdverbreid worden beschouwd, laat minder overlap zien in de kenmerkenruimten tussen en binnen klimaatzones (figuur 5C). Hoewel xerische bossen nu geclusterd zijn met de rest van de tropische biomen, vertonen deze habitats samen met tropisch grasland grote overlap in functionele ruimte met gematigde biomen zoals mediterrane bossen en gematigde graslanden.
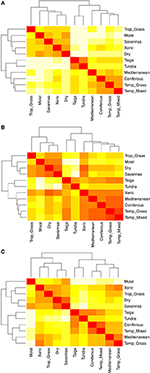
Figuur 5. (A) Paarsgewijze verschillen in soortensamenstelling tussen biomen. (B) Paarsgewijze verschillen in kenmerkhypervolumes (1-Sørensen similarity) tussen biomen op basis van het totale aantal soorten. (C) Paarsgewijze ongelijksoortigheid in kenmerk-hypervolumes (1-Sørensen similariteit) tussen biomen, waarbij alleen gebruik wordt gemaakt van de soorten die als functioneel vergelijkbaar en wijdverbreid worden beschouwd. Hoe lichter de cel, hoe groter de ongelijksoortigheid.
Discussie
Onze analyses leveren drie belangrijke inzichten op voor het begrijpen van terrestrische biomen door een functionele lens. Ten eerste tonen we aan dat, ondanks de vooruitgang in de compilatie en synthese van primaire biodiversiteitsgegevens, er nog steeds aanzienlijke kennistekorten bestaan die ons vermogen kunnen beperken om de functionele biodiversiteit van biomen op continentale tot wereldwijde schaal te kwantificeren. Ten tweede blijkt uit onze analyses van de beschikbare gegevens niettemin dat alle biomen in Noord- en Zuid-Amerika een opmerkelijk gemeenschappelijk patroon vertonen, waarbij de soorten die geografisch het meest verbreid zijn in een biome, de neiging vertonen gemeenschappelijke functionele kenmerken te delen, terwijl de soorten die functioneel het meest onderscheidend zijn, steevast beperkt zijn in hun verspreiding. Ten derde, wanneer alleen de wijdverspreide en functioneel gemeenschappelijke soorten worden beschouwd, kunnen biomen gemakkelijker functioneel worden onderscheiden, en patronen van ongelijksoortigheid tussen biomen blijken een overeenkomst te weerspiegelen tussen klimaat en de functionele nicheruimte van planten. Alles bij elkaar suggereren onze resultaten dat, hoewel de studie van de functionele diversiteit van biomen zich nog in de beginfase bevindt, de verdere ontwikkeling van het veld waarschijnlijk inzichten zal opleveren die evolutie, biogeografie, gemeenschapsopbouw en ecosysteemfunctie met elkaar verbinden.
Kennislacunes
De BIEN-database is specifiek ontworpen om lacunes in onze kennis van plantaardige biodiversiteit te dichten, maar zoals Hortal et al. (2015) aangeven, leiden interacterende kennistekorten tot onzekerheid bij het kwantificeren van biodiversiteit op de grootste schalen. Zo had niet alleen minder dan 10% van onze in kaart gebrachte soorten zowel kenmerkgegevens als fylogenetische informatie beschikbaar, maar die ontbrekende kenmerkgegevens (~66% van de kenmerkwaarden van soorten) waren behoorlijk gestructureerd, zowel fylogenetisch als geografisch. Het Raunkiaeriaanse tekort aan functionele kenmerkgegevens (sensu Hortal et al., 2015) interageert dus met het Wallaceaanse tekort aan distributie-informatie en het Darwiniaanse tekort aan fylogenetisch inzicht. En hoewel we fylogenetische kennis kunnen gebruiken om die ontbrekende waarden redelijk te imputeren (Swenson et al., 2017), blijven verschillende aspecten van de resulterende patronen die belangrijk zijn voor de indeling van biomen onzeker. De bestaande groeivormgegevens suggereren bijvoorbeeld dat houtachtige soorten domineren in alle tropische biomen, terwijl de proportionele diversiteit van kruidachtige groeivormen veel hoger is in gematigde en polaire biomen (zie ook Engemann et al., 2016; Zanne et al., 2018). Is deze latitudinale toename in relatieve diversiteit van kruidachtige planten een weerspiegeling van steekproefbiases en een gebrek aan taxonomische kennis over tropische kruidachtige planten, of weerspiegelt het de verschillende evolutionaire en biogeografische geschiedenissen in de Nearctische en Neotropische rijken? Dominante groeivormen zijn een van de belangrijkste onderscheidende kenmerken van biomen, dus systematische veranderingen in de meest diverse groeivormen (dominant of niet) zullen noodzakelijkerwijs van invloed zijn op onze voorspellingen over hoe de functionele diversiteit in biomen zou kunnen veranderen, en hoe die veranderingen de ecosysteemfunctie en -diensten zullen beïnvloeden.
We gebruikten verspreidingskaarten van soorten als invoergegevens, voornamelijk om het onbetwistbare probleem van vertekening door steekproeven te ondervangen, dat kenmerkend is voor datasets die uit meerdere bronnen zijn samengesteld (zie aanvullende figuur 1). Bereikkingskaarten afkomstig van verspreidingsmodellen van soorten kunnen echter een extra element van onzekerheid in onze resultaten vormen. Bijvoorbeeld, gezien de geografische resolutie van deze studie en de ruimtelijke complexiteit van bepaalde biomen, zouden deze modellen de geografische reikwijdte van sommige soorten kunnen hebben overschat van cellen van een enkel biome tot cellen van nabijgelegen en aan elkaar grenzende biomen. Het gebruik van gegevens over het voorkomen van soorten heeft de algemene resultaten van deze studie niet veranderd (zie de aanvullende figuren 2-6); een geringe afname van de functionele overlap tussen biomen zou echter wel degelijk een overschatting van het verspreidingsgebied van sommige soorten aan het licht kunnen brengen (zie de aanvullende figuren 2, 5, en 6). Het gebruik van informatie over het voorkomen heeft ook de algemene relaties tussen functioneel onderscheidend vermogen en wijdverspreidheid in onze resultaten veranderd, als gevolg van de relatieve overbemonstering van gematigde vs. tropische gebieden (zie aanvullende figuur 4). In dit geval wordt onze maat voor wijdverspreidheid beperkt door de uiterst beperkte bemonstering van de meeste soorten in tropische biomen. Naarmate de datasets en de bemonstering verbeteren, zal er waarschijnlijk ruimte zijn om deze onzekerheden in de toekomst te verminderen.
Het andere gebrek aan gegevens dat moet worden aangepakt om de functionele diversiteit van biomen te begrijpen, is het Prestoniaanse tekort aan informatie over abundantie. Abundantie is bijzonder belangrijk in studies van functionele diversiteit, omdat de eigenschappen die het meest van belang zijn voor het samenstellen van gemeenschappen, het functioneren van ecosystemen en biogeochemische cycli, die van de meest dominante soorten zijn. Omdat we onze assemblages baseerden op verspreidingskaarten afgeleid van verspreidingsmodellen voor soorten, konden we ons alleen richten op de geografische component van algemeenheid, maar de positieve relatie tussen lokale abundantie en geografisch bereik, de zogenaamde occupancy-abundance relatie, suggereert dat de meest wijdverspreide soorten in elk biome vaak ook tot de meer overvloedige soorten behoren (Borregaard en Rahbek, 2010; Novosolov et al., 2017). Omdat biomen over het algemeen worden gekenmerkt door de dominante groeivormen in de regio, zou het integreren van abundantie-informatie kunnen resulteren in biomen die minder functionele overlap vertonen. Maar hoewel BIEN omvangrijke plotgegevens heeft verzameld die schattingen van lokale abundantie mogelijk maken, wordt het maken van redelijke vergelijkingen tussen verschillende regio’s en groeivormen belemmerd door ongelijke bemonstering, regionale verschillen in gammadiversiteit, en incommensurate methoden om abundantie te kwantificeren.
Gemeenschappelijke patronen binnen biomen
Ondanks de aanhoudende hiaten in de beschikbare gegevens, brachten onze analyses een voorheen niet gedocumenteerde relatie aan het licht die alle biomen van het westelijk halfrond gemeen hebben: in elk biome hebben de meest verbreide soorten ook de neiging een laag functioneel onderscheidend vermogen te vertonen, terwijl de functioneel meest onderscheidende soorten bijna altijd beperkt zijn in hun verspreiding (Figuur 3). Deze “bezettingsgraad-redundantie” relatie kan suggereren dat de klimatologische omstandigheden die heersen binnen een biome selecteren op een set van gemeenschappelijke kenmerken, waarbij meer functioneel onderscheidende soorten beperkt zijn tot een zeldzamere subset van habitats binnen het biome. De algemene prevalentie van functioneel vergelijkbare soorten in alle biomen en de “bezetting-redundantie” relatie zijn beide consistent met een recente wereldwijde analyse van functionele diversiteit op gemeenschapsniveau die suggereert dat habitatfiltering leidt tot het naast elkaar bestaan van functioneel vergelijkbare soorten (Li et al., 2018), evenals studies die aantonen dat functionele redundanties de stabiliteit van gemeenschappen vergroten (bijv. Walker et al., 1999; Pillar et al., 2013). Bovendien, samen met de hoge mate van zowel taxonomische als functionele overlap tussen biomen (figuren 1, 5), versterkt het feit dat veel voorkomende, wijdverspreide soorten functioneel vergelijkbaar zijn de notie dat landplanten in een breed scala van milieuomstandigheden gemeenschappelijke kenmerken delen in de buurt van de kern van het functionele kenmerkenspectrum (Wright et al., 2004; Díaz et al., 2016). In tegenstelling tot Umaña et al. (2017), laten onze resultaten zien dat zeldzame, geografisch beperkte soorten al dan niet functioneel verschillend zijn van meer wijdverspreide en algemene soorten. De vraag blijft dus open of meer functioneel onderscheidende soorten specialisten zijn in bepaalde milieus of dat hun eigenschappencombinaties resulteren in demografische of fysiologische trade-offs die hun geografische verspreiding beperken.
Vergelijkingen tussen biomen
Onze vergelijking van eigenschappenhypervolumedistributies tussen biomen (Figuur 2B) is consistent met de waarneming dat soortenrijkere milieus ook functioneel diverser zijn (Swenson et al, 2012; Lamanna et al., 2014; Li et al., 2018; Šímová et al., 2018). Onze resultaten zijn meer dubbelzinnig met betrekking tot de hypothese dat seizoensgebonden en extreme klimatologische omgevingen de functionele diversiteit van soorten consistent beperken (de la Riva et al., 2018). Alle tropische biomen vertonen een hogere gemiddelde functionele diversiteit dan alle gematigde biomen, en de polaire biomen vertonen wel de kleinste hypervolumes. Echter, binnen elke groep vertonen drogere of meer seizoensvariabele biomen niet altijd kleinere hypervolumes (bijv. droge bossen, xerische bossen), zoals we zouden hebben verwacht volgens de tolerantiehypothese van Currie et al. (2004), terwijl optimale klimatologische omstandigheden meer combinaties van fysiologische parameters ondersteunen. Uit deze resultaten alleen kunnen we niet bepalen of gematigde en polaire biomen minder taxonomisch divers zijn vanwege beperkingen in de functionele niche-ruimte, of dat hun functionele hypervolumes klein zijn omdat ze niet taxonomisch divers zijn. Null-modeling benaderingen zouden mogelijk kunnen helpen om taxonomische en functionele diversiteit te ontwarren (Swenson et al., 2012; Lamanna et al., 2014; Šímová et al., 2018), maar een dergelijke analyse viel buiten het bestek van deze studie. Belangrijker is dat onze resultaten het belang versterken van inzicht in hoe evolutionaire en biogeografische geschiedenis vorm geven aan de functionele diversiteit van biomen (Woodward et al., 2004; Pennington et al., 2006; Donoghue en Edwards, 2014; Moncrieff et al., 2016). De uitgebreide uitwisseling van soorten (en hogere lineages) tussen biomen binnen, maar grotendeels niet tussen, verschillende biogeografische rijken zou zowel kunnen dienen om de functionele diversiteit binnen rijken te homogeniseren als om aanwijzingen te geven over de karakteristieke eigenschappen die door verschillende milieus worden geselecteerd of juist tegen geselecteerd (Douma et al, 2012; Zanne et al., 2014, 2018).
Ondanks substantiële hypervolume overlap tussen alle biomen (Supplementary Figure 2), blijken tropische, gematigde en koude biomen allemaal onderscheidbare regio’s van functionele ruimte te bezetten (Figuren 5B,C). De belangrijkste kenmerken die biomen van elkaar onderscheiden lijken kenmerken te zijn die gerelateerd zijn aan de totale omvang van de plant, met inbegrip van zowel de volwassen hoogte als de zaadmassa (Aanvullende Figuren 7-10), in plaats van kenmerken met betrekking tot de bladeconomie, zoals waargenomen in meer lokale, plot-gebaseerde analyses (bijv. Douma et al., 2012). De uitzondering is blad P, dat aanzienlijke verschillen vertoonde tussen tropische en gematigde/polaire biomen (Supplementary Figures 7-10), een patroon dat is waargenomen in andere analyses op basis van zowel soortspecifieke waarden als metingen van het gehele ecosysteem (Kerkhoff et al., 2005; Swenson et al., 2012). Omdat blad P ons meest dun bemonsterde kenmerk was, zouden de verschillen tussen tropische en gematigde/polaire gebieden onderhevig kunnen zijn aan vertekeningen van de imputatieprocedure. Blad P vertoont echter een significant fylogenetisch signaal (Kerkhoff et al., 2006), wat suggereert dat de imputaties onvertekend zouden moeten zijn (Swenson et al., 2017). Blad P is ook significant hoger in kruidachtige groeivormen (Kerkhoff et al., 2006), dus de latitudinale verschuiving van overwegend houtachtige naar kruidachtige diversiteit (Figuur 4) kan ook van invloed zijn op dit patroon.
Wanneer we hypervolume analyses beperkten tot alleen de meest wijdverspreide en functioneel meest voorkomende soorten in elk biome, overlapten de afzonderlijke biomen binnen elk biogeografisch rijk minder in functionele eigenschappenruimte (vergelijk Figuren 5B,C), wat suggereert dat deze soorten fenotypen kunnen weerspiegelen die beter zijn aangepast aan bepaalde omgevingen. In deze context is de xerische bosbiom bijzonder interessant. Gedeeltelijk als gevolg van de opname van de Chaco ecoregio, zijn xerische bossen taxonomisch geclusterd met de tropische biomen (Figuur 5A). Maar wanneer de functionele hypervolumes van alle soorten in aanmerking worden genomen, vertonen ze een veel sterkere gelijkenis met de gematigde biomen (figuur 5B). Tenslotte, wanneer alleen de meest verspreide en functioneel meest voorkomende soorten worden geanalyseerd, vertonen xerische bossen weer een grotere gelijkenis met tropische biomen, maar ook zij behouden een hoge mate van gelijkenis met gematigd grasland en mediterrane biomen (figuur 5C). Overgangen van warme, mesische omgevingen naar koudere, drogere en meer seizoensgebonden omgevingen worden vergemakkelijkt door vergelijkbare kenmerken, bijvoorbeeld kruidachtige habitus (Douma et al., 2012; Zanne et al., 2014, 2018), en net als koudere omgevingen hebben gemeenschappen in droge omgevingen ook de neiging om meer fylogenetisch geclusterd te zijn (Qian en Sandel, 2017). Bovendien maakt de positie van xerische zones op de grens van de inter-tropische convergentiezone ze tot een geografische overgang tussen het tropische en gematigde rijk. Het feit dat xerische bossen zowel functioneel als biogeografisch tussen de tropische en gematigde rijken in liggen, versterkt nog eens het idee dat we, om de functionele diversiteit van biomen beter te begrijpen, rekening moeten houden met hun biogeografische en fylogenetische geschiedenis (Pennington et al., 2006; Donoghue en Edwards, 2014; Moncrieff et al., 2016).
Conclusies
Elke classificatie van terrestrische biomen legt een klein aantal discrete categorieën op aan continue gradiënten in klimaat en soortenverdelingen, en vormt dus een grove vereenvoudiging van het complexe ecologische landschap (Moncrieff et al., 2016, maar zie Silva de Miranda et al., 2018). Toch zijn biomen, ondanks hun potentieel voor oversimplificatie, nuttige constructen voor het organiseren en begrijpen van de biodiversiteit en het functioneren van de belangrijkste terrestrische ecosystemen van de aarde, en op eigenschappen gebaseerde benaderingen hebben een groot potentieel om te helpen bij het dynamisch modelleren van wereldwijde vegetatiedistributies.
In deze studie hebben we aangetoond dat een groot deel van de taxonomische diversiteit van alle biomen soorten vertegenwoordigt die zowel smal gedistribueerd als functioneel vergelijkbaar zijn. Verder hebben binnen biomen de meest functioneel onderscheidende soorten ook de neiging om geografisch zeldzaam te zijn, terwijl wijdverspreide soorten uniform een laag functioneel onderscheidend vermogen vertonen. Ondanks uitgebreide taxonomische en functionele overlap tussen de biomen, clusteren ze in onderscheidbare, biogeografisch en klimatologisch verschillende eenheden, vooral wanneer de functionele clustering gebaseerd is op de meest verspreide en functioneel meest voorkomende soorten in elk biome. Het bevorderen van een functioneel begrip van biomen vereist echter niet alleen een betere karakterisering van de variatie in eigenschappen binnen en tussen vegetatietypen, maar ook informatie over de biogeografische en fylogenetische geschiedenis van soortenassemblages en de relatieve abundantie van soorten binnen biomen.
Beschikbaarheid van gegevens
Trait datasets en kaarten van het soortenbereik die voor deze studie zijn geanalyseerd, kunnen worden gedownload via het R-pakket BIEN (Maitner et al., 2018). Zie http://bien.nceas.ucsb.edu/bien/ voor meer details over de BIEN-database.
Bijdragen van auteurs
SE-L, AK, BJE, DMN, en CV ontwierpen de studie; SE-L en AK analyseerden de gegevens; SE-L en AK schreven de eerste versie van het manuscript; CV, BB, NK, BSM, BM, RKP, BS, SS, J-CS, en SKW droegen bij aan de collatie en creatie van de traits en range maps database. Alle auteurs hebben bijgedragen aan de ontwikkeling en het schrijven van het manuscript.
Funding
SE-L, DMN, en AK werden gesteund door een collaborative research grant van de US National Science Foundation (DEB-1556651). BJE werd gesteund door National Science Foundation award DEB-1457812 en Macrosystems-1065861. CV werd ondersteund door de European Research Council (Grant ERC-StG-2014-639706-CONSTRAINTS) en door de French Foundation for Research on Biodiversity (FRB; www.fondationbiodiversite.fr) in het kader van het CESAB-project Causes and consequences of functional rarity from local to global scales. J-CS beschouwt dit werk als een bijdrage aan zijn VILLUM Investigator-project Biodiversity Dynamics in a Changing World, gefinancierd door VILLUM FONDEN (subsidie 16549). SKW werd ondersteund door het Strategic Science Investment Fund van de Science and Innovation Group van het Nieuw-Zeelandse ministerie van Handel, Innovatie en Werkgelegenheid.
Conflict of Interest Statement
De auteurs verklaren dat het onderzoek is uitgevoerd in afwezigheid van commerciële of financiële relaties die zouden kunnen worden opgevat als een potentieel belangenconflict.
Acknowledgments
Wij danken Dan Griffith en twee reviewers voor hun opbouwende kritiek die dit manuscript sterk heeft verbeterd. Wij danken ook alle BIEN-databijdragers (zie http://bien.nceas.ucsb.edu/bien/data-contributors/ voor een volledige lijst).
Aanvullend materiaal
Het aanvullend materiaal voor dit artikel is online te vinden op: https://www.frontiersin.org/articles/10.3389/fevo.2018.00219/full#supplementary-material
Banda, R.K., Delgado-Salinas, A., Dexter, K. G., Linares-Palomino, R., Oliveira-Filho, A., Prado, D., et al. (2016). Plantendiversiteitspatronen in neotropische droge bossen en hun implicaties voor het behoud. Science 353, 1383-1387. doi: 10.1126/science.aaf5080
PubMed Abstract | CrossRef Full Text | Google Scholar
Blonder, B., Lamanna, C., Violle, C., and Enquist, B. J. (2014). Het n-dimensionale hypervolume. Glob. Ecol. Biogeogr. 23, 595-609. doi: 10.1111/geb.12146
CrossRef Full Text | Google Scholar
Blonder, B., Morrow, C. B., Maitner, B., Harris, D. J., Lamanna, C., Violle, C., et al. (2018). Nieuwe benaderingen voor het afbakenen van n-dimensionale hypervolumes. Methods Ecol. Evol. 9, 305-319. doi: 10.1111/2041-210X.12865
CrossRef Full Text | Google Scholar
Bonan, G. B., Oleson, K. W., Fisher, R. A., Lasslop, G., and Reichstein, M. (2012). Reconciling leaf physiological traits and canopy flux data: use of the TRY and FLUXNET databases in the Community Land Model version 4. J. Geophys. Res. Biogeosci. 117:G02026. doi: 10.1029/2011JG001913
CrossRef Full Text | Google Scholar
Borregaard, M. K., and Rahbek, C. (2010). Causaliteit van de relatie tussen geografische verspreiding en soortenrijkdom. Q. Rev. Biol. 85, 3-25. doi: 10.1086/650265
PubMed Abstract | CrossRef Full Text | Google Scholar
Bruggeman, J., Heringa, J., and Brandt, B. W. (2009). PhyloPars: schatting van ontbrekende parameterwaarden met behulp van fylogenie. Nucleic Acids Res. 37, W179-W184. doi: 10.1093/nar/gkp370
PubMed Abstract | CrossRef Full Text | Google Scholar
Cadotte, M. W., Carscadden, K., and Mirotchnick, N. (2011). Beyond species: functionele diversiteit en de instandhouding van ecologische processen en diensten. J. Appl. Ecol. 48, 1079-1087. doi: 10.1111/j.1365-2664.2011.02048.x
CrossRef Full Text | Google Scholar
Cornwell, W. K., Cornelissen, J. H., Amatangelo, K., Dorrepaal, E., Eviner, V. T., Godoy, O., et al. (2008). Plantensoortkenmerken zijn de belangrijkste determinant van de afbraaksnelheid van strooisel in biomen wereldwijd. Ecol. Lett. 11, 1065-1071. doi: 10.1111/j.1461-0248.2008.01219.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Craine, J. M. (2009). Resource Strategies of Wild Plants. New Jersey, NJ: Princeton University Press. doi: 10.1515/9781400830640
CrossRef Full Text | Google Scholar
Currie, D. J., Mittelbach, G. G., Cornell, H. V., Field, R., Guégan, J. F., Hawkins, B. A., et al. (2004). Predictions and tests of climate-based hypotheses of broad-scale variation in taxonomic richness. Ecol. Lett. 12, 1121-1134. doi: 10.1111/j.1461-0248.2004.00671.x
CrossRef Full Text | Google Scholar
de la Riva, E. G., Violle, C., Pérez-Ramos, I. M., Marañón, T., Navarro-Fernández, C. M., Olmo, M., et al. (2018). A multidimensional functional trait approach reveals the imprint of environmental stress in Mediterranean woody Communities. Ecosystems 21, 248-262. doi: 10.1007/s10021-017-0147-7
CrossRef Full Text | Google Scholar
Díaz, S., Kattge, J., Cornelissen, J. H., Wright, I. J., Lavorel, S., Dray, S., et al. (2016). Het globale spectrum van plantenvorm en -functie. Nature 529, 167-171. doi: 10.1038/nature16489
PubMed Abstract | CrossRef Full Text | Google Scholar
Díaz, S., Lavorel, S., de Bello, F., Quétier, F., Grigulis, K., and Robson, T. M. (2007). Incorporating plant functional diversity effects in ecosystem service assessments. Proc. Natl. Acad. Sci. U.S.A 104, 20684-20689. doi: 10.1073/pnas.0704716104
PubMed Abstract | CrossRef Full Text | Google Scholar
Donoghue, M. J., and Edwards, E. J. (2014). Biome shifts and niche evolution in plants. Annu. Rev. Ecol. Evol. Syst. 45, 547-572. doi: 10.1146/annurev-ecolsys-120213-091905
CrossRef Full Text | Google Scholar
Douma, J. C., Aerts, R., Witte, J. P. M., Bekker, R. M., Kunzmann, D., Metselaar, K., et al. (2012). Een combinatie van functioneel verschillende plantenkenmerken biedt een middel om kwantitatief een breed scala van soortenassemblages in NW Europa te voorspellen. Ecography 35, 364-373. doi: 10.1111/j.1600-0587.2011.07068.x
CrossRef Full Text | Google Scholar
Engemann, K., Sandel, B., Enquist, B. J., Jørgensen, P. M., Kraft, N., Marcuse-Kubitza, A., et al. (2016). Patterns and drivers of plant functional group dominance across the Western Hemisphere: a macroecological re-assessment based on a massive botanical dataset. Bot. J. Linn. Soc. 180, 141-160. doi: 10.1111/boj.12362
CrossRef Full Text | Google Scholar
Enquist, B. J., Condit, R., Peet, R., Thiers, B., and Schildhauer, M. (2016). Het botanische informatie- en ecologienetwerk (BIEN): cyberinfrastructuur voor een geïntegreerd botanisch informatienetwerk om de ecologische effecten van wereldwijde klimaatverandering op de biodiversiteit van planten te onderzoeken. PeerJ Prepr. 4:e2615v2. doi: 10.7287/peerj.preprints.2615v2
CrossRef Full Text | Google Scholar
Garnier, E., Cortez, J., Billès, G., Navas, M. L., Roumet, C., Debussche, M., et al. (2004). Plant functional markers capture ecosystem properties during secondary succession. Ecology 85, 2630-2637. doi: 10.1890/03-0799
CrossRef Full Text | Google Scholar
Goldsmith, G. R., Morueta-Holme, N., Sandel, B., Fitz, E. D., Fitz, S. D., Boyle, B., et al. (2016). Plant-O-Matic: een dynamische en mobiele gids voor alle planten van de Amerika’s. Methods Ecol. Evol. 7, 960-965. doi: 10.1111/2041-210X.12548
CrossRef Full Text | Google Scholar
Goolsby, E. W., Bruggeman, J., and Ané, C. (2017). Rphylopars: fast multivariate phylogenetic comparative methods for missing data and within-species variation. Methods Ecol. Evol. 8, 22-27. doi: 10.1111/2041-210X.12612
CrossRef Full Text | Google Scholar
Grenié, M., Denelle, P., Tucker, C. M., Munoz, F., and Violle, C. (2017). funrar: an R package to characterize functional rarity. Divers. Distrib. 23, 1365-1371. doi: 10.1111/ddi.12629
CrossRef Full Text | Google Scholar
Hortal, J., de Bello, F., Diniz-Filho, J. A. F., Lewinsohn, T. M., Lobo, J. M., and Ladle, R. J. (2015). Seven shortfalls that beset large-scale knowledge of biodiversity. Annu. Rev. Ecol. Evol. Syst. 46, 523-549. doi: 10.1146/annurev-ecolsys-112414-054400
CrossRef Full Text | Google Scholar
Kerkhoff, A. J., Enquist, B. J., Elser, J. J., and Fagan, W. F. (2005). Plant allometry, stoichiometry and the temperature-dependence of primary productivity. Glob. Ecol. Biogeogr. 14, 585-598. doi: 10.1111/j.1466-822X.2005.00187.x
CrossRef Full Text | Google Scholar
Kerkhoff, A. J., Fagan, W. F., Elser, J. J., and Enquist, B. J. (2006). Fylogenetic and growth form variation in the scaling of nitrogen and phosphorus in the seed plants. Am. Nat. 168, E103-E122. doi: 10.1086/507879
PubMed Abstract | CrossRef Full Text | Google Scholar
Kraft, N. J., Valencia, R., and Ackerly, D. D. (2008). Functional traits and niche-based tree community assembly in an Amazonian forest. Science 322, 580-582. doi: 10.1126/science.1160662
PubMed Abstract | CrossRef Full Text | Google Scholar
Lamanna, C., Blonder, B., Violle, C., Kraft, N. J., Sandel, B., et al. (2014). Functional trait space and the latitudinal diversity gradient. Proc. Natl. Acad. Sci. U.S.A. 111, 13745-13750. doi: 10.1073/pnas.1317722111
PubMed Abstract | CrossRef Full Text | Google Scholar
Lavorel, S., and Garnier, E. (2002). Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the Holy Grail. Funct. Ecol. 16, 545-556. doi: 10.1046/j.1365-2435.2002.00664.x
CrossRef Full Text | Google Scholar
Lavorel, S., Grigulis, K., Lamarque, P., Colace, M.-P., Garden, D., Girel, J., et al. (2011). Het gebruik van functionele eigenschappen van planten om de landschapsdistributie van meervoudige ecosysteemdiensten te begrijpen. J. Ecol. 99, 135-147. doi: 10.1111/j.1365-2745.2010.01753.x
CrossRef Full Text | Google Scholar
Li, Y., Shipley, B., Price, J. N., Dantas, V. de L., Tamme, R., Westoby, M., et al. (2018). Habitatfiltering bepaalt de functionele nichebezetting van plantengemeenschappen wereldwijd. J. Ecol. 106, 1001-1009. doi: 10.1111/1365-2745.12802
CrossRef Full Text | Google Scholar
Magallón, S., Gómez-Acevedo, S., Sánchez-Reyes, L. L., and Hernández-Hernández, T. (2015). A metacalibrated time-tree documents the early rise of flowering plant phylogenetic diversity. New Phytol. 207, 437-453. doi: 10.1111/nph.13264
PubMed Abstract | CrossRef Full Text | Google Scholar
Maitner, B. S., Boyle, B., Casler, N., Condit, R., Donoghue, J., Durán, S. M., et al. (2018). Het BIEN R pakket: een tool om toegang te krijgen tot de botanische informatie en ecologie netwerk (BIEN) database. Methods Ecol. Evol. 9, 373-379. doi: 10.1111/2041-210X.12861
CrossRef Full Text | Google Scholar
Moncrieff, G. R., Bond, W. J., and Higgins, S. I. (2016). Revising the biome concept for understanding and predicting global change impacts. J. Biogeogr. 43, 863-873. doi: 10.1111/jbi.12701
CrossRef Full Text | Google Scholar
Novosolov, M., Rodda, G. H., North, A. C., Butchart, S. H. M., Tallowin, O. J. S., Gainsbury, A. M., et al. (2017). Population density-range size relationship revisited. Glob. Ecol. Biogeogr. 26, 1088-1097. doi: 10.1111/geb.12617
CrossRef Full Text | Google Scholar
Olson, D. M., Dinerstein, E., Wikramanayake, E. D., Burgess, N. D., Powell, G. V. N., Underwood, E. C., et al. (2001). Terrestrial Ecoregions of the world: a new map of life on earth: een nieuwe wereldkaart van terrestrische ecoregio’s biedt een innovatief instrument voor het behoud van de biodiversiteit. Bioscience 51, 933-938.doi: 10.1641/0006-3568(2001)0512.0.CO;2
CrossRef Full Text | Google Scholar
Pennington, R. T., Prado, D. E., and Pendry, C. A. (2000). Neotropical seasonally dry forests and quaternary vegetation changes. J. Biogeogr. 27, 261-273. doi: 10.1046/j.1365-2699.2000.00397.x
CrossRef Full Text | Google Scholar
Pennington, R. T., Richardson, J. E., and Lavin, M. (2006). Inzichten in de historische constructie van soortenrijke biomen uit gedateerde plantenfylogenieën, neutrale ecologische theorie en fylogenetische gemeenschapsstructuur. New Phytol. 172, 605-616. doi: 10.1111/j.1469-8137.2006.01902.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Penone, C., Davidson, A. D., Shoemaker, K. T., Di Marco, M., Rondinini, C., Brooks, T. M., et al. (2014). Imputatie van ontbrekende data in life-history trait datasets: welke benadering presteert het beste? Methods Ecol. Evol. 5, 961-970. doi: 10.1111/2041-210X.12232
CrossRef Full Text | Google Scholar
Pillar, V. D., Blanco, C. C., Müller, S. C., Sosinski, E. E., Joner, F., and Duarte, L. D. S. (2013). Functionele redundantie en stabiliteit in plantengemeenschappen. J. Veg. Sci. 24, 963-974. doi: 10.1111/jvs.12047
CrossRef Full Text | Google Scholar
Prado, D. E., and Gibbs, P. E. (1993). Patterns of species distributions in the dry seasonal forests of South America. Ann. Mo. Bot. Gard. 80, 902-927. doi: 10.2307/2399937
CrossRef Full Text | Google Scholar
Qian, H., and Sandel, B. (2017). Fylogenetic structure of regional angiosperm assemblages across latitudinal and climatic gradients in North America. Glob. Ecol. Biogeogr. 26, 1258-1269. doi: 10.1111/geb.12634
CrossRef Full Text | Google Scholar
Reich, P. B., Wright, I. J., Cavender-Bares, J., Craine, J. M., Oleksyn, J., Westoby, M., et al. (2003). De evolutie van functionele variatie in planten: eigenschappen, spectra, en strategieën. Int. J. Plant Sci. 164, S143-S164. doi: 10.1086/374368
CrossRef Full Text | Google Scholar
Silva de Miranda, P. L., Oliveira-Filho, A., Pennington, R. T., Neves, D. M., Baker, T. R., and Dexter, K. G. (2018). Using tree species inventories to map biomes and assess their climatic overlaps in lowland tropical South America. Glob. Ecol. Biogeogr. 27, 899-912. doi: 10.1111/geb.12749
CrossRef Full Text | Google Scholar
Šímová, I., Violle, C., Kraft, N. J. B., Storch, D., Svenning, J.-C., Boyle, B., et al. (2015). Shifts in trait means and variances in North American tree assemblages: species richness patterns are loosely related to the functional space. Ecography 38, 649-658. doi: 10.1111/ecog.00867
CrossRef Full Text | Google Scholar
Šímová, I., Violle, C., Svenning, J.-C., Kattge, J., Engemann, K., Sandel, B., et al. (2018). Spatial patterns and climate relationships of major plant traits in the New World differ between woody and herbaceous species. J. Biogeogr. 45, 895-916. doi: 10.1111/jbi.13171
CrossRef Full Text | Google Scholar
Smith, S. A., and Beaulieu, J. M. (2009). Life history influences rates of climatic niche evolution in flowering plants. Proc. Biol. Sci. 276, 4345-4352. doi: 10.1098/rspb.2009.1176
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, S. A., and Brown, J. W. (2018). Constructing a broadly inclusive seed plant phylogeny. Am. J. Bot. 105, 302-314. doi: 10.1002/ajb2.1019
PubMed Abstract | CrossRef Full Text | Google Scholar
Smith, S. A., and Donoghue, M. J. (2008). Snelheden van moleculaire evolutie zijn gekoppeld aan levensgeschiedenis in bloeiende planten. Science 322, 86-89. doi: 10.1126/science.1163197
PubMed Abstract | CrossRef Full Text | Google Scholar
Swenson, N. G. (2014). Phylogenetic imputation of plant functional trait databases.Ecography 37, 105-110. doi: 10.1111/j.1600-0587.2013.00528.x
CrossRef Full Text
Swenson, N. G., Enquist, B. J., Pither, J., Kerkhoff, A. J., Boyle, B., Weiser, M. D., et al. (2012). De biogeografie en filtering van de functionele diversiteit van houtachtige planten in Noord- en Zuid-Amerika. Glob. Ecol. Biogeogr. 21, 798-808. doi: 10.1111/j.1466-8238.2011.00727.x
CrossRef Full Text | Google Scholar
Swenson, N. G., Weiser, M. D., Mao, L., Araújo, M. B., Diniz-Filho, J. A. F., Kollmann, J., et al. (2017). Fylogenie en de voorspelling van boom functionele diversiteit in nieuwe continentale settings. Glob. Ecol. Biogeogr. 26, 553-562. doi: 10.1111/geb.12559
CrossRef Full Text | Google Scholar
Umaña, M. N., Mi, X., Cao, M., Enquist, B. J., Hao, Z., Howe, R., et al. (2017). De rol van functionele uniciteit en ruimtelijke aggregatie in het verklaren van zeldzaamheid bij bomen. Glob. Ecol. Biogeogr. 26, 777-786. doi: 10.1111/geb.12583
CrossRef Full Text | Google Scholar
van Bodegom, P. M., Douma, J. C., and Verheijen, L. M. (2014). A fully traits-based approach to modeling global vegetation distribution. Proc. Natl. Acad. Sci. U.S.A. 111, 13733-13738. doi: 10.1073/pnas.1304551110
PubMed Abstract | CrossRef Full Text | Google Scholar
Violle, C., Reich, P. B., Pacala, S. W., Enquist, B. J., and Kattge, J. (2014). De opkomst en belofte van functionele biogeografie. Proc. Natl. Acad. Sci. U.S.A. 111, 13690-13696. doi: 10.1073/pnas.1415442111
PubMed Abstract | CrossRef Full Text | Google Scholar
Violle, C., Thuiller, W., Mouquet, N., Munoz, F., Kraft, N. J. B., Cadotte, M. W., et al. (2017). Functionele zeldzaamheid: de ecologie van uitschieters. Trends Ecol. Evol. 32, 356-367. doi: 10.1016/j.tree.2017.02.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Walker, B., Kinzig, A., and Langridge, J. (1999). Plant attribute diversity, resilience, and ecosystem function: the nature and significance of dominant and minor species. Ecosystems 2, 95-113. doi: 10.1007/s100219900062
CrossRef Full Text | Google Scholar
Westoby, M. (1998). A leaf-height-seed (LHS) plant ecology strategy scheme. Plant Soil 199, 213-227. doi: 10.1023/A:1004327224729
CrossRef Full Text | Google Scholar
Woodward, F. I., Lomas, M. R., and Kelly, C. K. (2004). Global climate and the distribution of plant biomes. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 359, 1465-1476. doi: 10.1098/rstb.2004.1525
PubMed Abstract | CrossRef Full Text | Google Scholar
Wright, I. J., Reich, P. B., Westoby, M., Ackerly, D. D., Baruch, Z., Bongers, F., et al. (2004). The worldwide leaf economics spectrum. Nature 428, 821-827. doi: 10.1038/nature02403
PubMed Abstract | CrossRef Full Text | Google Scholar
Xia, J., Niu, S., Ciais, P., Janssens, I. A., Chen, J., Ammann, C., et al. (2015). Gezamenlijke controle van terrestrische bruto primaire productiviteit door plantenfenologie en -fysiologie. Proc. Natl. Acad. Sci. U.S.A. 112, 2788-2793. doi: 10.1073/pnas.1413090112
PubMed Abstract | CrossRef Full Text | Google Scholar
Zanne, A. E., Pearse, W. D., Cornwell, W. K., McGlinn, D. J., Wright, I. J., and Uyeda, J. C. (2018). Functionele biogeografie van angiospermen: leven aan de uitersten. New Phytol. 218, 1697-1709. doi: 10.1111/nph.15114
PubMed Abstract | CrossRef Full Text | Google Scholar
Zanne, A. E., Tank, D. C., Cornwell, W. K., Eastman, J. M., Smith, S. A., FitzJohn, R. G., et al. (2014). Drie sleutels tot de radiatie van angiospermen in vriesomgevingen. Nature 506, 89-92. doi: 10.1038/nature12872
PubMed Abstract | CrossRef Full Text | Google Scholar
.