Assessment | Biopsychology | Comparative |Cognitive | Developmental | Language | Individual differences |Personality | Philosophy | Social |
Methods | Statistics |Clinical | Educational | Industrial |Professional items |World psychology |
Biological:Gedragsgenetica – Evolutionaire psychologie – Neuroanatomie – Neurochemie – Neuroendocrinologie -Neurowetenschappen – Psychoneuroimmunologie – Fysiologische Psychologie – Psychofarmacologie(Index, Outline)
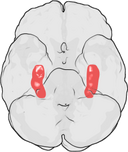
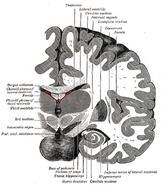
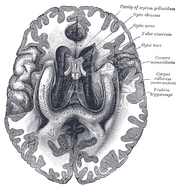
De plaats van de hippocampus in het menselijk brein.
Voor het gelijknamige tijdschrift, zie Hippocampus (tijdschrift)
De hippocampus is een deel van de voorhersenen, gelegen in de mediale temporale kwab. Het behoort tot het limbisch systeem en speelt een belangrijke rol bij het langetermijngeheugen en de ruimtelijke navigatie. Mensen en andere zoogdieren hebben twee hippocampi, één aan elke kant van de hersenen. Bij knaagdieren, waar hij het meest uitgebreid is bestudeerd, heeft de hippocampus de vorm van een banaan. Bij de mens heeft hij een gebogen en kronkelige vorm die vroege anatomisten aan een zeepaardje deed denken. De naam is in feite afgeleid van het Griekse woord voor zeepaardje (Grieks: hippos = paard, campos = zee).
Bij de ziekte van Alzheimer is de hippocampus een van de eerste gebieden in de hersenen die schade oplopen; geheugenproblemen en desoriëntatie behoren tot de eerste symptomen. Schade aan de hippocampus kan ook het gevolg zijn van zuurstofgebrek (anoxie), encefalitis of mesiale temporale kwab epilepsie. Bij mensen met uitgebreide schade aan de hippocampus kan amnesie optreden, d.w.z. het onvermogen om nieuwe herinneringen te vormen of vast te houden.
- Functies van de hippocampus
- Rol in het geheugen
- Rol in het ruimtelijk geheugen en navigatie
- Hippocampus en depressie
- Geschiedenis
- Anatomie
- Physiology
- Het theta ritme
- Scherpe golven
- Evolutie
- Zie ook
- Referenties & Bibliografie
- Sleutelteksten
- Boeken
- Papers
- Aanvullend materiaal
- Papers
- Extra afbeeldingen
Functies van de hippocampus
Het eerste idee was misschien dat de hippocampus betrokken is bij de reukzin: dit lijkt vooral te zijn gesuggereerd door de plaats ervan in de hersenen, naast de reukcortex. Er bestaat nog steeds enige belangstelling voor olfactorische reacties van de hippocampus, maar bijna niemand gelooft nu dat de voornaamste functie van de hippocampus olfactorisch is.
In de loop der jaren hebben drie hoofdideeën over de functie van de hippocampus de literatuur gedomineerd: inhibitie, geheugen, en ruimte. De gedragsremmingstheorie (door O’Keefe en Nadel gekarikaturiseerd als “stap op de rem!”) was tot in de jaren zestig zeer populair. Het ontleende veel van zijn kracht aan twee observaties: ten eerste, dieren met hippocampus schade hebben de neiging hyperactief te zijn; ten tweede, dieren met hippocampus schade hebben vaak moeite om te leren om reacties te remmen die hen eerder zijn aangeleerd. Jeffrey Gray ontwikkelde deze gedachtegang tot een volwaardige theorie over de rol van de hippocampus bij angst. De inhibitietheorie is momenteel echter niet erg populair.
De tweede belangrijke gedachtelijn houdt verband met de hippocampus en het geheugen. Hoewel er voorlopers waren, ontleende dit idee zijn voornaamste kracht aan een zeer bekend verslag van Scoville en Milner over de resultaten van chirurgische vernietiging van de hippocampus (in een poging om epileptische aanvallen te verlichten), bij een patiënt bekend als H.M. Het onverwachte resultaat was ernstig geheugenverlies: H.M. was niet in staat zich bewust gebeurtenissen te herinneren die plaatsvonden na zijn operatie of gedurende verscheidene jaren daarvoor. Dit geval wekte zo’n enorme belangstelling op dat H.M. nu het meest intensief bestudeerde medische geval in de geschiedenis zou zijn. In de daaropvolgende jaren zijn ook andere patiënten met vergelijkbare niveaus van hippocampusschade en geheugenverlies (veroorzaakt door een ongeval of ziekte) bestudeerd, en letterlijk duizenden experimenten hebben de fysiologie van neurale plasticiteit in de hippocampus bestudeerd. Men is het er thans vrijwel algemeen over eens dat de hippocampus een belangrijke rol speelt in het geheugen; over de precieze aard van deze rol wordt echter nog uitvoerig gediscussieerd.
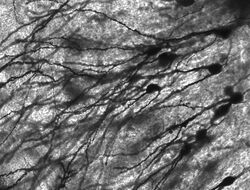
Golgi-gekleurde neuronen in de knaagdierhippocampus.
Rol in het geheugen
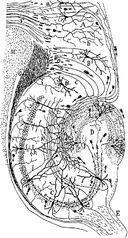
Tekening van het neurale circuit van de knaagdierhippocampus. S. Ramón y Cajal, 1911.
Psychologen en neurowetenschappers betwisten de precieze rol van de hippocampus, maar zijn het er in het algemeen over eens dat deze een essentiële rol speelt bij de vorming van nieuwe herinneringen aan persoonlijk beleefde gebeurtenissen (episodisch of autobiografisch geheugen). Sommige onderzoekers geven er de voorkeur aan de hippocampus te beschouwen als een deel van een groter mediaal temporaal kwab geheugensysteem dat verantwoordelijk is voor het algemeen declaratief geheugen (herinneringen die expliciet verbaal kunnen worden vastgelegd – dit zou bijvoorbeeld het geheugen voor feiten omvatten naast het episodisch geheugen).
Hoofdartikel: Rol van de hippocampus in het geheugen
Er zijn aanwijzingen dat de hippocampus betrokken is bij het opslaan en verwerken van ruimtelijke informatie. Studies bij ratten hebben aangetoond dat neuronen in de hippocampus een ruimtelijk ontstekingsveld hebben. Deze cellen worden plaatscellen genoemd. Sommige cellen vuren wanneer het dier zich op een bepaalde plaats bevindt, ongeacht de reisrichting, terwijl de meeste tenminste gedeeltelijk gevoelig zijn voor de richting van de kop en de reisrichting.
Hoofdartikel: Rol van de hippocampus in het ruimtelijk geheugen en navigatie
Hippocampus en depressie
- Hippocampus volumeverlies, wellicht een weerspiegeling van de effecten van overmatige glucocorticoïden op
neurogenese, is vastgesteld bij groepen depressieve mensen in vergelijking met controles.
Hoofdartikel: Depressie en de hippocampus
Geschiedenis
De anatoom Giulio Cesare Aranzi (circa 1564) gebruikte voor het eerst de term hippocampus om het hersenorgaan aan te duiden vanwege de visuele gelijkenis met een zeepaardje. Dit orgaan werd aanvankelijk in verband gebracht met de reukzin, eerder dan met zijn gekende functie in het verwerven van geheugen. De Rus Vladimir Bekhterev merkte de rol van de hippocampus in het geheugen op rond 1900, op basis van observaties van een patiënt met ernstige geheugenstoornissen. Jarenlang was de conventionele opvatting over de hippocampus echter dat deze, net als de rest van het limbisch systeem, verantwoordelijk was voor emoties.
Het belang van de hippocampus voor het geheugen werd onder de aandacht van onderzoekers gebracht door patiënt HM. HM leed aan een aantal anterograde en temporally-grade retrograde geheugenstoornissen (dergelijke stoornissen zijn het onderwerp van de film Memento) na de bilaterale verwijdering van verschillende mediale-temporale kwab structuren (met inbegrip van bilaterale ablatie van zijn hippocampi) om frequente epileptische aanvallen te verlichten. Van bijzonder belang is dat HM was nog steeds in staat om procedurele taken (die worden geassocieerd met het striatum) te leren en had een bovengemiddeld IQ. HM toonde een opvallende single-dissociatie tussen intelligentie en declaratief geheugen. De relatieve omvang van de hippocampusvorming in verhouding tot het totale volume van de hersenen is bij de meeste zoogdiersoorten vaak geconserveerd. Niettemin is vastgesteld dat deze gebieden bij walvisachtigen relatief hypotroof zijn.
Anatomie
Hoofdartikel: Anatomie van de hippocampus
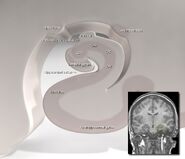
Humane hippocampus.
Nissl-gekleurde coronale doorsnede van de hersenen van een makaakaap, waarop de hippocampus (omcirkeld) te zien is. Bron: brainmaps.org
Anatomisch gezien is de hippocampus een uitwerking van de rand van de cortex. Hij kan worden onderscheiden als een zone waar de cortex zich vernauwt tot een enkele laag van zeer dicht opeengepakte neuronen, die zich omkrult in een strakke S-vorm. De structuren die langs de rand van de cortex liggen, vormen het zogenaamde limbisch systeem (Latijn limbus = grens): deze omvatten de hippocampus, de cingulate cortex, de olfactorische cortex en de amygdala. Paul MacLean suggereerde ooit, als onderdeel van zijn drievoudige hersentheorie, dat de limbische structuren de neurale basis van emotie omvatten. De meeste neurowetenschappers geloven echter niet langer dat het concept van een verenigd “limbisch systeem” geldig is.
De hippocampus, als geheel, lijkt uiteindelijk op een gebogen buis, die op verschillende manieren is geanaloogd met een zeepaardje, of een ramshoorn (Cornu Ammonis), of een banaan.Deze algemene lay-out geldt voor het hele scala van zoogdiersoorten, van egel tot mens, hoewel de details variëren. Bij de rat lijken de twee hippocampi verbazend veel op een paar bananen, verbonden bij de stam. Bij de mens en de aap is het gedeelte van de hippocampus onderaan, bij de basis van de temporale kwab, veel breder dan het gedeelte bovenaan. Een van de gevolgen van deze complexe geometrie is dat dwarsdoorsneden door de hippocampus een verbijsterende variëteit aan vormen kunnen vertonen, afhankelijk van de hoek en de plaats van de snede.
De sterkste verbindingen van de hippocampus zijn met de entorhinale cortex (EC), die ernaast ligt in de temporale kwab. De oppervlakkige lagen van de EC leveren de meeste inputs aan de hippocampus, en de diepe lagen van de EC ontvangen de meeste outputs. De EC is op zijn beurt sterk, en wederzijds, verbonden met vele andere delen van de cortex. De hippocampus ontvangt ook een zeer belangrijke projectie van het mediale septale gebied. Vernietiging van het septale gebied doet het theta-ritme in de hippocampus verdwijnen en brengt ernstige schade toe aan bepaalde vormen van geheugen. (Van de zogenaamde “date rape” drugs wordt verondersteld dat zij hun amnestische effecten ten minste gedeeltelijk uitoefenen door de cholinerge projectie van het mediale septum naar de hippocampus tegen te werken.)
Physiology
Voorbeelden van ratten hippocampale EEG en CA1 neurale activiteit in de theta (wakker/gedrag) en LIA (slow-wave slaap) modes. Elke plot toont 20 seconden van gegevens, met een hippocampale EEG spoor op de top, spike rasters van 40 gelijktijdig opgenomen CA1 piramidale cellen in het midden (elk raster lijn vertegenwoordigt een andere cel), en een plot van de loopsnelheid aan de onderkant. De bovenste grafiek representeert een periode waarin de rat actief op zoek was naar verspreide voedselbolletjes. In de onderste grafiek sliep de rat.
De hippocampus vertoont twee belangrijke “modes” van activiteit, elk geassocieerd met een verschillend patroon van EEG-golven en neurale populatie-activiteit. Deze modes zijn genoemd naar de EEG-patronen die ermee geassocieerd zijn: theta en grote onregelmatige activiteit (LIA). Hier volgen enkele van hun voornaamste kenmerken bij de rat, het dier dat het meest uitvoerig is bestudeerd:
De theta modus verschijnt tijdens staten van actief, alert gedrag (vooral voortbeweging), en ook tijdens REM (dromen) slaap. In de theta-modus wordt het EEG gedomineerd door grote regelmatige golven met een frequentiebereik van 6-9 Hz, en de belangrijkste groepen hippocampale neuronen (piramidale cellen en granule cellen) vertonen spaarzame populatie-activiteit, wat betekent dat in elk kort tijdsinterval, de overgrote meerderheid van de cellen stil zijn, terwijl de kleine overblijvende fractie vuren met een relatief hoge snelheid, tot 50 spikes in een seconde voor de meest actieve van hen. Een actieve cel blijft typisch actief gedurende een halve seconde tot enkele seconden. Naarmate de rat zich gedraagt, vallen de actieve cellen stil en worden nieuwe cellen actief, maar het totale percentage actieve cellen blijft min of meer constant. In veel situaties wordt de cellactiviteit grotendeels bepaald door de ruimtelijke locatie van het dier, maar ook andere gedragsvariabelen zijn duidelijk van invloed.
De LIA modus verschijnt tijdens de slow-wave (niet-dromende) slaap, en ook tijdens staten van wakkere onbeweeglijkheid, zoals rusten of eten. In de LIA modus, wordt het EEG gedomineerd door scherpe golven, die willekeurig getimede grote ombuigingen van het EEG-signaal gedurende 200-300msec. Deze scherpe golven bepalen ook het neurale activiteitspatroon van de populatie. Daartussen zijn de piramidale cellen en de granule cellen zeer rustig (maar niet stil). Tijdens een scherpe golf kan 5-10% van de populatie actiepotentialen uitzenden gedurende een periode van 50 msec; veel van deze cellen zenden niet één maar een uitbarsting van spikes uit.
Deze twee hippocapampale activiteitsmodi kunnen zowel bij primaten als bij ratten worden waargenomen, met de belangrijke uitzondering dat het moeilijk is gebleken een robuuste theta-ritmiek waar te nemen in de primatenhippocampus. Er zijn echter kwalitatief vergelijkbare scherpe golven, en vergelijkbare toestand-afhankelijke veranderingen in neurale populatie-activiteit.
Het theta ritme
Omwille van zijn dicht opeengepakte neurale lagen, genereert de hippocampus enkele van de grootste EEG-signalen van elke hersenstructuur. In sommige situaties wordt het EEG gedomineerd door regelmatige golven, die vaak vele seconden aanhouden. Dit EEG-patroon staat bekend als het theta-ritme. Het was een van de vroegste EEG-verschijnselen die werden ontdekt: de eerste beschrijving kwam van Jung en Kornmuller, in 1938. Het was echter pas in 1954, met de publicatie door Green en Arduini van een lange en grondige studie van het theta-ritme bij konijnen, katten en apen, dat de belangstelling echt op gang kwam. Misschien grotendeels omdat zij het theta-ritme in verband brachten met opwinding, het onderwerp van die tijd, lokte hun artikel een vloedgolf van vervolgstudies uit, die resulteerden in de publicatie van letterlijk honderden studies over de fysiologie en farmacologie van theta in de jaren vijftig en zestig van de vorige eeuw. Ondanks deze ontzagwekkende hoeveelheid werk, bleven vele vragen onbeantwoord, vooral de vraag naar de functie. Zelfs op dit ogenblik is deze meest kritische vraag nog niet overtuigend beantwoord.
Theta ritmiciteit is zeer duidelijk bij konijnen en knaagdieren, en ook duidelijk aanwezig bij katten en honden. Of theta kan worden waargenomen bij primaten is een lastige vraag. Green en Arduini rapporteerden slechts zeer korte uitbarstingen van tamelijk onregelmatige ritmiek bij apen, en de meeste latere studies hebben weinig meer gezien. Verschillen in methodologie hebben het echter moeilijk gemaakt om sterke conclusies te trekken.
In ratten (de dieren die veruit het meest uitvoerig zijn bestudeerd), wordt theta voornamelijk gezien in twee omstandigheden: ten eerste, wanneer een dier loopt of op een andere manier actief in interactie is met zijn omgeving; ten tweede, tijdens de REM slaap. De frequentie neemt toe als functie van de loopsnelheid, beginnend bij ongeveer 6,5 Hz aan de lage kant en oplopend tot ongeveer 9 Hz aan de hoge kant, hoewel hogere frequenties soms worden waargenomen bij dramatische bewegingen zoals sprongen over grote tussenruimten. Bij andere, grotere, diersoorten liggen de theta-frequenties over het algemeen iets lager. De gedragsafhankelijkheid lijkt ook per soort te verschillen: bij katten en konijnen wordt theta vaak waargenomen tijdens een toestand van onbeweeglijke alertheid. Dit is ook waargenomen bij ratten, maar alleen wanneer zij hevig geschrokken zijn.
Theta is niet alleen beperkt tot de hippocampus. Bij ratten kan het worden waargenomen in vele delen van de hersenen, waaronder bijna alle delen die sterk interageren met de hippocampus. De pacemaker voor het ritme zou in het mediale septale gebied liggen: dit gebied projecteert op alle gebieden die theta ritmiciteit vertonen, en vernietiging ervan elimineert theta in de gehele hersenen. (Er kan een uitzondering zijn, een klein gebied in de hypothalamus genaamd de supramamillarynucleus, die in staat lijkt te zijn om theta onafhankelijk van het septum in sommige situaties te ondersteunen.)
De functie van theta, aangenomen dat het er een heeft, is nog niet overtuigend verklaard, hoewel er talrijke theorieën zijn voorgesteld.De meest populaire trend is om het te relateren aan leren en geheugen. Het is bekend dat letsels van het mediale septum – het centrale knooppunt van het theta systeem – ernstige verstoringen van het geheugen veroorzaken. Het mediale septum is echter meer dan alleen de regelaar van theta, het is ook de belangrijkste bron van cholinerge projecties naar dehippocampus. Het is niet vastgesteld dat septumlaesies hun effecten specifiek uitoefenen door theta uit te schakelen.
Scherpe golven
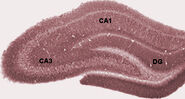
Tijdens de slaap, of tijdens waaktoestanden wanneer een dier rust of op een andere manier niet bezig is met zijn omgeving, vertoont het hippocampale EEG een patroon van onregelmatige trage golven, iets groter in amplitude dan theta-golven. Dit patroon wordt nu en dan onderbroken door grote pieken, de zogenaamde scherpe golven. Deze gebeurtenissen gaan gepaard met uitbarstingen van piekactiviteit, met een duur van 50-100 msec, in piramidale cellen van CA3 enCA1. Zij worden ook geassocieerd met kortdurende hoogfrequente EEG-oscillaties die “rimpelingen” worden genoemd. Rimpelingen, met frequenties in het bereik 150-200 Hz bij ratten, kunnen gewoonlijk alleen worden gedetecteerd door elektroden die zich in of zeer dicht bij de CA1-cellichaamlaag bevinden. Daarentegen zullen elektroden die zich overal in de hippocampus bevinden, of zelfs in naburige hersenstructuren, vaak scherpe golven oppikken als grote trage EEG-afwijkingen, die 200-400 msec duren.
In ratten zijn scherpe golven het meest robuust tijdens de slaap, wanneer zij voorkomen met een gemiddelde snelheid van ongeveer 1 per seconde, maar in een zeer onregelmatig temporaal patroon. Scherpe golven komen ook voor tijdens de inactieve waaktoestand, maar zij zijn dan minder frequent en gewoonlijk kleiner. Scherpe golven zijn ook waargenomen in de temporale kwab van de mens en in de hippocampus van de aap. Bij apen zijn scherpe golven vrij robuust, maar ze komen lang niet zo vaak voor als bij ratten.
Een van de interessantste aspecten van scherpe golven is dat ze geassocieerd lijken te zijn met het geheugen. Wilson en McNaughton 1994, en tal van latere studies, gemeld dat wanneer hippocampal plaatscellen overlappende ruimtelijke vuren velden (en dus vaak vuren in de buurt van-simultaneity) hebben, ze de neiging om gecorreleerde activiteit vertonen tijdens de slaap na de gedragssessie. Deze toename van correlatie, algemeen bekend als reactivatie, blijkt voornamelijk beperkt te zijn tot scherpe golven. Er is voorgesteld dat scherpe golven in feite reactiveringen zijn van neurale activiteitspatronen die tijdens het gedrag werden onthouden, aangedreven door de versterking van synaptische verbindingen binnen de hippocampus. Dit idee vormt een belangrijk onderdeel van de “twee fasen geheugen” theorie, bepleit door Buzsaki en anderen, die stelt dat herinneringen worden opgeslagen in de hippocampus tijdens het gedrag, en later overgebracht naar de neocortex tijdens de slaap: scherpe golven worden gesuggereerd om Hebbische synaptische veranderingen in daneocorticale doelen van hippocampus output pathways rijden.
Evolutie
De hippocampus ziet er over het algemeen ongeveer hetzelfde uit bij alle zoogdiersoorten, van basale zoals de egel tot de meest “geavanceerde” zoals de mens. De verhouding hippocampusgrootte/lichaamsgrootte neemt in het algemeen toe en is ongeveer twee keer zo groot bij primaten als bij de egel. De hippocampus neemt bij knaagdieren dus een veel groter deel van de corticale massa in beslag dan bij primaten.
Er is ook een algemeen verband tussen de grootte van de hippocampus en het ruimtelijk geheugen: bij vergelijkingen tussen soorten van dezelfde soort hebben soorten met een groter ruimtelijk geheugen doorgaans een groter hippocampusvolume. Deze relatie strekt zich ook uit tot sekseverschillen: in soorten waar mannetjes en vrouwtjes sterke verschillen vertonen in ruimtelijk geheugen, vertonen ze ook vaak overeenkomstige verschillen in het volume van de hippocampus
Niet-zoogdiersoorten hebben geen hersenstructuur die lijkt op de hippocampus van zoogdieren, maar ze hebben er wel een die als homoloog wordt beschouwd. De hippocampus is, zoals gezegd, in wezen de mediale rand van de cortex. Alleen zoogdieren hebben een volledig ontwikkelde cortex, maar de structuur waaruit deze is geëvolueerd, het pallium genaamd, is aanwezig bij alle gewervelde dieren, zelfs bij de meest primitieve, zoals de lamprei of de slijmprik. Het pallium wordt gewoonlijk verdeeld in drie zones: mediaal, lateraal en dorsaal. Het mediale pallium vormt de voorloper van de hippocampus. Het lijkt visueel niet op de hippocampus, omdat de lagen niet in een S-vorm zijn gebogen en ook niet omwikkeld zijn door de dentate gyrus, maar de homologie blijkt uit sterke chemische en functionele affiniteiten. Er zijn nu aanwijzingen dat deze hippocampus-achtige structuren betrokken zijn bij het ruimtelijk inzicht bij vogels, reptielen en vissen.
Bij vogels is de overeenkomst zo duidelijk dat de meeste anatomisten naar de mediale palliale zone verwijzen als de “aviaire hippocampus”. Talrijke vogelsoorten hebben een sterk ruimtelijk inzicht, vooral vogels die voedsel opslaan. Er zijn aanwijzingen dat vogels die voedsel bewaren een grotere hippocampus hebben dan andere vogelsoorten en dat schade aan de hippocampus het ruimtelijk geheugen aantast.
Het verhaal voor vissen is complexer. Bij teleostvissen (die de grote meerderheid van de bestaande soorten vormen), is de voorhersenen vreemd vervormd in vergelijking met andere soorten gewervelde dieren. De meeste neuroanalogen menen dat de voorhersenen van de teleostvis in wezen omgekeerd zijn, als een binnenstebuiten gekeerde sok, zodat structuren die bij de meeste gewervelde dieren binnenin, naast de ventrikels liggen, bij teleostvissen aan de buitenkant te vinden zijn, en omgekeerd. Een van de gevolgen hiervan is dat het mediale pallium (“hippocampus”-zone) van een typisch gewerveld dier wordt geacht overeen te komen met het laterale pallium van een typisch vis. Bij verschillende soorten vissen (met name goudvissen) is experimenteel aangetoond dat zij een sterk ruimtelijk geheugen hebben en zelfs “cognitieve kaarten” vormen van de gebieden waar zij leven. Er zijn bewijzen dat beschadiging van het laterale pallium het ruimtelijk geheugen aantast. (Lange-afstandsnavigatie, zoals homing door zalmen, lijkt echter op andere mechanismen te berusten.)
Dus lijkt de rol van de hippocampus in de navigatie ver terug in de evolutie van de gewervelde dieren te beginnen, vóór de splitsingen die honderden miljoenen jaren geleden plaatsvonden. Het is nog niet bekend of het mediale pallium een soortgelijke rol speelt bij nog primitievere gewervelde dieren, zoals haaien en roggen, of zelfs lampreien en slijmprikken. Sommige soorten insecten en weekdieren zoals de octopus hebben ook een sterk ruimtelijk leer- en navigatievermogen, maar deze blijken anders te werken dan het ruimtelijk systeem van de zoogdieren, zodat er nog geen goede reden is om te denken dat zij een gemeenschappelijke evolutionaire oorsprong hebben; evenmin is er voldoende gelijkenis in hersenstructuur om bij deze soorten iets te kunnen identificeren dat lijkt op een “hippocampus”.
Zie ook
- Amygdalohippocampectomie
- Mediane voorhersenenbundel
- Septale kernen
Referenties & Bibliografie
Sleutelteksten
Boeken
- Aboitiz, F, Morales D, Montiel J (2003). De evolutionaire oorsprong van de isocortex van zoogdieren: Naar een geïntegreerde ontwikkelings- en functionele benadering. Behav. Brain Sciences 26: 535-552.
- Amaral, D; Lavenex P (2006). “Ch 3. Hippocampal Neuroanatomy” Andersen P, Morris R, Amaral D, Bliss T, O’Keefe J The Hippocampus Book, Oxford University Press.
- Broglio, C, Gómez A, Durán E, Ocaña FM, Jiménez-Moya F, Rodríguez F, Salas C (2002). Kenmerken van een gemeenschappelijk plan voor de voorhersenen van vertebraten: Gespecialiseerde palliale gebieden voor ruimtelijk, temporeel en emotioneel geheugen bij actinopterygische vissen. Brain Res. Bull. 57: 397-399.
- Buzsáki, G (2002). Theta oscillaties in de hippocampus. Neuron 33: 325-340.
- Buzsáki, G (2006). Rhythms of the Brain.
- Cantero, JL, Atienza M, Stickgold R, Kahana MJ, Madsen JR, Kocsis B (2003). Slaap-afhankelijke theta oscillaties in de menselijke hippocampus en neocortex. J. Neurosci. 23: 10897-10903.
- Colombo, M, Broadbent N (2000). Is de aviaire hippocampus een functionele homoloog van de hippocampus bij zoogdieren? Neurosci. Biobehav. Rev. 24: 465-484.
- Eichnbaum, H; Cohen NJ (1993). Memory, Amnesia, and the Hippocampal System, MIT Press.
- Ekstrom, AD, Kahana MJ, Caplan JB, Fields TA, Isham EA, Newman EL, Fried I (2003). Cellulaire netwerken die ten grondslag liggen aan de menselijke ruimtelijke navigatie. Nature 425: 184-188.
- Gray, JA; McNaughton N (2000). De neuropsychologie van angst: An Enquiry into the Functions of the Septo-Hippocampal System, Oxford University Press.
- Green, JD, Arduini AA (1954). Hippocampus elektrische activiteit in arousal. J. Neurophysiol. 17: 533-557.
- Jacobs, LF, Gaulin SJ, Sherry DF, Hoffman GE (1990). Evolution of spatial cognition: sex-specific patterns of spatial behavior predict hippocampal size. PNAS 87: 6349-6352.
- Jacobs, LF (2003). De evolutie van de cognitieve kaart. Brain Behav. Evol. 62: 128-139.
- Kirk, IJ, McNaughton N (1991). Supramammillary cell firing and hippocampal rhythmical slow activity. Neuroreport 11: 723-725.
- Maguire, EA, Gadian DG, Johnsrude IS, Good CD, Ashburner J, Frackowiak RS, Frith CD (2000). Navigatie-gerelateerde structurele verandering in de hippocampi van taxichauffeurs. PNAS 97: 4398-4403.
- McNaughton, BL, Battaglia FP, Jensen O, Moser EI, Moser MB (2006). Padintegratie en de neurale basis van de ‘cognitieve kaart’. Nat. Rev. Neurosci. 7: 663-678.
- Moser, EI, Kropf E, Moser M-B (2008). Place Cells, Grid Cells, and the Brain’s Spatial Representation System (Plaatscellen, rastercellen en het ruimtelijk representatiesysteem van de hersenen). Ann. Rev. Neurosci. 31.
- Nieuwenhuys, R (1982). Een overzicht van de organisatie van de hersenen van Actinopterygische Vissen. Am. Zool. 22: 287-310.
- O’Kane, G, Kensinger EA, Corkin S (2004). Bewijs voor semantisch leren bij diepe amnesie: Een onderzoek met patiënt H.M.. Hippocampus 14: 417-425.
- O’Keefe, J; Nadel L (1978). The Hippocampus as a Cognitive Map, Oxford University Press.
- Portavella, M, Vargas JP, Torres B, Salas C (2002). The effects of telencephalic pallial lesions on spatial, temporal, and emotional learning in goldfish. Brain Res. Bull. 57: 397-399.
- Rodríguez, F, Lópeza JC, Vargasa JP, Broglioa C, Gómeza Y, Salas C (2002). Spatial memory and hippocampal pallium through vertebrate evolution: insights from reptiles and teleost fish. Brain Res. Bull. 57: 499-503.
- Sainsbury, RS, Heynen A, Montoya CP (1987). Behavioral correlates of hippocampal type 2 theta in the rat. Physiol. Behav. 39: 513-519.
- Scoville, WB, Milner B (1957). Loss of Recent Memory After Bilateral Hippocampal Lesions. J. Neurol. Neurosurg. Psych. 20: 11-21.
- Shettleworth, SJ (2003). Geheugen en hippocampale specialisatie bij voedselproducerende vogels: Uitdagingen voor onderzoek naar vergelijkende cognitie. Brain Behav. Evol. 62: 108-116.
- Skaggs, WE, McNaughton BL, Wilson MA, Barnes CA (1996). Theta fase precessie in hippocampale neuronale populaties en de compressie van temporele sequenties. Hippocampus 6: 149-176.
- Squire, LR (1992). Memory and the hippocampus: a synthesis from findings with rats, monkeys, and humans. Psych. Rev. 99: 195-231.
- Squire, LR; Schacter DL (2002). The Neuropsychology of Memory, Guilford Press.
- Vanderwolf, CH (1969). Hippocampale elektrische activiteit en vrijwillige beweging bij de rat. EEG & Clin. Neurophysiol. 26: 407-418.
- Vargas, JP, Bingman VP, Portavella M, López JC (2006). Telencephalon en geometrische ruimte bij goudvissen. Eur. J. Neurosci. 24: 2870-2878.
- West, MJ (1990). Stereological studies of the hippocampus: a comparison of the hippocampal subdivisions of various species including hedgehogs, laboratory rodents, wild mice and men. Prog. Brain Res. 83: 13-36.
Papers
- Amaral DG and Cowan WM. 1980. Subcortical afferents to the hippocampal formation in the monkey. Journal of Comparative Neurology. Feb 15; 189(4):573-91.
- Duvernoy, H. (2005) The Human Hippocampus, 3rd ed. Berlin: Springer-Verlag.
Aanvullend materiaal
Papers
- Google Scholar
- http://www.psycheducation.org/emotion/hippocampus.htm Great brain tours!!!
- BrainMaps at UCDavis hippocampus
- Artificial Hippocampus
- BrainInfo at the University of Washington hier-164
- John O’Keefe & Lynn Nadel (1978) The Hippocampus as a Cognitive Map , Oxford University Press. Full Text
Extra afbeeldingen
Een foto aan deze galerij toevoegen
Hersenen van de mens: Limbisch systeem
Amygdala – Cingulate gyrus – Fornicate gyrus – Hippocampus – Hypothalamus – Mammillary body – Nucleus accumbens – Orbitofrontale cortex – Parahippocampale gyrus
|
v-d-e
Hersenen van de mens: voorhersenen (cerebrum – hersenschors – hersenhelften, grijze stof) (TA A14.1.09.002-240, 301-320, GA 9.818-826) |
|||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Frontale kwab |
|
||||||||||||||||
| Pariëtale kwab |
|
Superolaterale
|
Mediale/inferieure
|
Beide
|
|||||||||||||
| Occipitale kwab |
|
Superolaterale
|
Mediaal/inferior
|
||||||||||||||
| Temporale kwab |
|
Superolateraal
|
Medial/inferior
|
||||||||||||||
| Interlobar sulci/fissures |
|
Superolateraal
|
Mediaal/inferieur
|
||||||||||||||
| Limbische kwab |
|
Anderen
|
|||||||||||||||
| Insulaire kwab |
Lange gyrus van insula – Korte gyri van insula – Cirkelvormige sulcus van insula |
||||||||||||||||
| Algemeen |
Operculum – polen van hersenhelften |
||||||||||||||||
Sommige categoriseringen zijn benaderingen, en sommige Brodmann-gebieden omvatten gyri.
|
|||||||||||||||||
Deze pagina maakt gebruik van Creative Commons-gelicentieerde inhoud van Wikipedia (bekijk auteurs).